Курс лекций
по дисциплине «Биохимия сельскохозяйственной продукции»
(для бакалавров технологического факультета по направлению: 35.03.07 -
Технология производства и переработки сельскохозяйственной
продукции)
Тверь, 2017
Составитель: к. с.-х. н., доцент кафедры агрохимии и земледелия
Шилова О.В.
Рецензент: к. б. н., доцент кафедры биологии животных, зоотехнии и основ
ветеринарии Тверской ГСХА Корецкая Е.А.
Шилова О.В. Курс лекций по дисциплине «Биохимия сельскохозяйственной продукции» (для бакалавров технологического факультета по направлению: 35.03.07 - Технология производства и переработки сельскохозяйственной
продукции) – Тверь, 2017. – 152 с.
Председатель методической комиссии  Акимов А.А.
Акимов А.А.
Введение
Цели дисциплины: формирование современных представлений, знаний и умений о превращениях веществ и энергии в живых организмах, химическом составе сельскохозяйственной продукции растительного и животного происхождения, биохимических процессах, происходящих в ней при хранении и переработке.
Задачи дисциплины: изучение строения и биологических функций важнейших органических веществ; механизмов ферментативных и биоэнергетических превращений в организмах; химического состава сельскохозяйственной продукции и биохимических процессов, происходящих в ней при хранении и переработке;
– оценка качества и технологических свойств сельскохозяйственной продукции по биохимическим показателям;
– применение знаний о химическом составе и биохимических процессах при обосновании технологий производства, хранения и переработки сельскохозяйственной продукции;
– ознакомление с современными методами и достижениями биохимической науки.
Модуль 1. Характеристика и биологические функции
Органических соединений
Тема 1. Введение в биохимию (ОПК-6)
План:
1. Определение биохимии как науки.
2. Важнейшие результаты биохимических исследований в первой половине 20-го века, позволившие сформулировать молекулярные концепции жизнедеятельности различных организмов.
3. Основные направления развития современной биохимической науки.
4. Применение достижений биохимии в промышленности, медицине, сельском хозяйстве.
Важнейшие результаты биохимических исследований в первой
Половине 20-го века, позволившие сформулировать молекулярные концепции жизнедеятельности различных организмов.
Только перечень фундаментальных наук химического направления уже говорит об исключительном разнообразии проявления химической формы движения материи и влиянии ее на нашу повседневную жизнь.
Иммуноферментный анализ, возникший более пятнадцати лет назад на пересечении иммунохимии и инженерной энзимологии, стал в настоящее время одним из распространенных методов исследования. Явные преимущества нового метода, к которым относится простота выполнения, доступность и стабильность реагентов, экспрессность и возможность автоматизации для проведения массовых анализов, обеспечили его прочное положение в клинической биохимии, при диагностике заболеваний растений и животных, в научных исследованиях. Благодаря успехам биотехнологии иммуноферментный анализ далее интенсивно развивался, поскольку с помощью генной инженерии были получены в высокоочищенном виде малодоступные антигены, а также ферменты-маркеры и их конъюгаты с антигенами, а с помощью клеточной инженерии — моноклональные антитела с заданной специфичностью и аффинностью. Новые направления развития иммуноферментного анализа связаны с использованием различных методов регуляции ферментативной активности при детектировании комплексов антиген — антитело.
Тема 2. Состав, строение и биологические функции углеводов
И липидов (ОПК-6)
План:
1. Общая характеристика и классификация углеводов.
2. Содержание сахаров и полисахаридов в сельскохозяйственной продукции. 3. Основные разновидности липидов и их значение для растений, животных и человека.
4. Содержание липидов в продуктах растительного, животного и микробного происхождения.
5. Строение и функции простых липидов – жира и воска.
Содержание сахаров и полисахаридов в сельскохозяйственной
Продукции.
Полисахариды - полимеры, мономерами которых служат моносахариды гексозы. Наиболее известными из дисахаридов (два мономера) являются сахароза и лактоза. Важнейшими полисахаридами являются крахмал и гликоген, служащие запасными веществами клеток растений и животных, а также целлюлоза - важнейший структурный компонент растительных клеток.
Растения обладают большим разнообразием углеводов, чем животные, так как способны синтезировать их на свету в процессе фотосинтеза. Важнейшие функции углеводов в клетке: энергетическая, структурная и запасающая.
Энергетическая роль состоит в том, что углеводы служат источником энергии в растительных и животных клетках; структурная — клеточная стенка у растений почти полностью состоит из полисахарида целлюлозы; запасающая - крахмал служит запасным продуктом растений. Он накапливается в процессе фотосинтеза в вегетационный период и у ряда растений откладывается в клубнях, луковицах и т. д. В животных клетках эту роль выполняет гликоген, откладывающийся преимущественно в печени.
В питании человека и животных углеводы составляют основную массу пищи. За счет углеводов обеспечивается 1/2 суточной энергетической потребности пищевого рациона человека. Углеводы способствуют предохранению белка от трат на энергетические цели. В сутки взрослому человеку необходимо 400-500 г углеводов (в том числе крахмала – 350-400г, сахаров – 50-100 г, других углеводов – 25 г), которые должны поступать с пищевыми
продуктами. При тяжелой физической нагрузке потребность в углеводах возрастает. При избыточном введении в организм человека углеводы могут превращаться в жиры или откладываться в небольших количествах в печени и мышцах в виде животного крахмала - гликогена.
С точки зрения пищевой ценности углеводы подразделяются на усваиваемые и неусваиваемые. Усваиваемые углеводы – моно и дисахариды, крахмал, гликоген. Неусваиваемые – целлюлоза, гемицеллюлозы, инулин, пектин, гумми, слизи.
В пищеварительном тракте человека усваиваемые углеводы (за исключением моносахаридов) расщепляются под действием ферментов до моносахаридов, которые через стенки кишечника всасываются в кровь и разносятся по всему телу.
При избытке простых углеводов и отсутствии расхода энергии часть углеводов превращается в жир или откладывается в печени как запасной источник энергии на временное хранение в виде гликогена.
Неусваиваемые углеводы организмом человека не утилизируются, но они чрезвычайно важны для пищеварения и составляют так называемые «пищевые волокна». Пищевые волокна стимулируют моторную функцию кишечника, препятствуют всасыванию холестерина, играют положительную роль в нормализации состава микрофлоры кишечника, в ингибировании гнилостных процессов, способствуют выведению из организма токсичных элементов. Суточная норма пищевых волокон составляет 20-25 г.
Животные продукты содержат мало углеводов, поэтому основным источником углеводов для человека служит растительная пища. Углеводы составляют три четверти сухой массы растений и водорослей, они содержатся в зерновых, фруктах, овощах. В растениях углеводы накапливаются как запасные вещества (например, крахмал) или они грают роль опорного материала (клетчатка).
Содержание углеводов, г на 100 г продукта (крахмал, сахара, неусвояемые углеводы, всего): мука пшеничная - 67-68; 1,7-1,8; 0,1-0,2; 73-74; макароны - 62-69; 1,7-4,6; 0,1-0,2; 72-75; рис – 55; 3; 4-10; 63-64; гречка - 63-64; 2; 1-2; 67-68; кукуруза – 57; 2,5-3,0; 6-10; 67-70; яблоки - 0,2; 14,90; 1,7; 11-17; персики - 15,8; 1,6; 17-18; виноград - 15,6-24,0; 1,2; 17-25; лимоны - 2,1; 1,6; 3-4; земляника - 6,5; 3,0; 9-10; картофель - 14-25.
Главными усваиваемыми углеводами в питании человека являются крахмал и сахароза. На долю крахмала приходится примерно 80 % всех потребляемых человеком углеводов. Крахмал является главным энергетические ресурсом человека.
Источники крахмала – зерновые, бобовые, картофель. Моносахариды и олигосахариды присутствуют в зерновых в относительно малых количествах. Сахароза обычно поступает в организм человека с продуктами, в которые она добавляется (кондитерские изделия, напитки, мороженое).
Продукты с высоким содержанием сахара являются наименее ценными из всех углеводных продуктов. Известно, что необходимо увеличивать содержание в рационе пищевых волокон. Источником пищевых волокон являются ржаные и пшеничные отруби, овощи, фрукты. Хлеб из цельного зерна с точки зрения содержания пищевых волокон гораздо более ценен, чем хлеб из муки высших сортов.
Углеводы плодов представлены в основном сахарозой, глюкозой, фруктозой, а также клетчаткой и пектиновыми веществами. Имеются продукты, состоящие почти из одних углеводов: крахмал, сахар, мед, карамель.
Животные продукты содержат значительно меньше углеводов, чем растительные. Одним из главнейших представителей животных крахмалов является гликоген. Мясной и печеночный гликоген своим строением похожи на крахмал. В молоке содержится лактоза: 4,7 % - в коровьем, 6,7 % - в человеческом.
Свойства углеводов и их превращения имеют большое значение при хранении и производстве пищевых продуктов. Так, во время хранения плодов и овощей происходит потеря массы в результате расхода углеводов на процессы дыхания. Превращения пектиновых веществ обуславливают изменение консистенции плодов.
Содержание липидов в продуктах растительного, животного и
Микробного происхождения.
Степень ненасыщенности и длина цепей высших карбоновых кислот (т. е. число атомов углерода) определяют физические свойства того или иного жира.
Жиры с короткими и непредельными кислотными цепями имеют низкую температуру плавления. При комнатной температуре это жидкости (масла) либо мазеподобные вещества (жиры). И наоборот, жиры с длинными и насыщенными цепями высших карбоновых кислот при комнатной температуре становятся твердыми. Вот почему при гидрировании (насыщении кислотных цепей атомами водорода по двойным связям) жидкое арахисовое масло, например, становится мазеобразным, а подсолнечное масло превращается в твердый маргарин. По сравнению с обитателями южных широт в организме животных, обитающих в холодном климате (например, у рыб арктических морей), обычно содержится больше ненасыщенных триацилглицеролов. По этой причине тело их остается гибким и при низких температурах.
Липиды могут образовывать комплексы с другими биологическими молекулами - белками и сахарами.
Липиды, или резервные жиры, представляют собой сложные молекулы, которые обычно называют жирными телами. Их принято классифицировать в соответствии с происхождением:
— липиды животного происхождения: к ним принадлежат жиры, содержащиеся в мясе, рыбе, масле, молочных продуктах и сыре, яйцах и т.д.
— липиды растительного происхождения: они содержатся в маслах (оливковом, подсолнечном и т.д.) и маргарине.
Однако, следовало бы их классифицировать по их химической формуле. Тогда жиры можно подразделить на два вида:
— насыщенные жирные кислоты, найденные в мясе, продуктах свинины, яйцах, молочных продуктах (молоко, масло, сливки, сыры), пальмовом масле и т.д.;
— мононенасыщенные и полиненасыщенные жирные кислоты, которые представляют из себя жиры, остающиеся жидкими при комнатной температуре (подсолнечное масло, оливковое масло, рапсовое масло и т.д.). Некоторые из них могут затвердевать при гидрогенизации (например, производство маргарина);
— ненасыщенные жиры, содержащиеся в рыбе, утке и гусе.
Потребление липидов, содержащихся в продуктах питания, является чрезвычайно важным поскольку:
— они снабжают организм энергией, накапливаемой в виде запасов жира и доступной при необходимости для снабжения тела глюкозой;
— они служат основой для формирования мембран и клеток;
— они входят в состав ткани и, в частности, нервной системы;
— они помогают производить гормоны и простагланди-ны;
— они формируют основу производства солей желчи;
— они содержат витамины растворимые в жирах, А, Д, Е,иК;
— они единственные источники того, что мы называем необходимыми жирными кислотами: линолевая и альфа-линоленовая кислоты;
— некоторые жирные кислоты способствуют предотвращению сердечно-сосудистых заболеваний.
а. Липиды и ожирение
Жиры — самые большие поставщики энергии, вот почему они главные враги в теории низкокалорийных диет. Они нарушают обмен веществ и ведут к образованию запасов жира. Повышенное содержание сахара в крови, возникшее из-за избытка инсулина, приводит к накоплению жировых излишков из липидов (липогенезис).
б. Липиды и холестерин
Взаимосвязь между излишним потреблением жиров и уровнем холестерина в крови (который является ответственным за сердечно-сосудистые заболевания) фактически показана. Целесообразно подразделять холестерин на два типа: хороший и плохой. В идеале следовало бы поддерживать общий уровень холестерина ниже или равным 2 граммам на литр и с как можно более высоким содержанием хорошего холестерина.
Помните, что не все жиры повышают плохой холестерин. Напротив, некоторые даже имеют тенденцию значительно снижать их. Этот вопрос мы рассмотрим более подробно в главе, посвященной повышенному содержанию холестерина в крови и опасности сердечно-сосудистых заболеваний.
С этой точки зрения следует классифицировать жиры на три новые категории.
Жиры, которые повышают холестерин. Это — насыщенные жирные кислоты, которые преимущественно содержатся в мясе, продуктах свинины, молоке и всех молочных продуктах, масле и некоторых сырах.
Излишнее потребление насыщенных жиров может привести к повышению уровня холестерина в крови и предрасположенности к сердечно-сосудистым заболеваниям. Многие исследователи склонны считать, что чрезмерное снисходительное отношение к насыщенным жирам приводит к опасности возникновения некоторых раковых образований.
Жиры, которые слабо влияют на холестерин - Это те, которые находятся в домашней птице, ракообразных и яйцах. Жиры, которые понижают холестерин и предотвращают артериальные проблемы
Это ненасыщенные жирные кислоты, которые преимущественно найдены в маслах (кроме пальмового масла), масляных семенах и рыбе, а также в гусином и утином жире (паштет из утиной печенки, и т.д.). Среди этих кислот нужно упомянуть следующие:
— Мононенасыщенные жирные кислоты, в особенности, олеиновая кислота из оливкового масла, которые обладают свойством снижения общего холестерина и увеличения хорошего холестерина. Их преимущество в химической устойчивости.
— Полиненасыщенные жирные кислоты, обнаруженные в подсолнечном масле, кукурузном и рапсовом маслах, понижают общий холестерин. Их много в необходимых жирных кислотах. Недостатком этих кислот является их легкая окисляемость. Эти полиненасыщенные жирные кислоты при окислении становятся такими же плохими для артерий как насыщенные жирные кислоты. Химические изменения, которым подвергаются некоторые растительные жиры, когда они затвердевают (становятся маргарином), похоже изменяют их свойства.
в. Необходимые жирные кислоты
Линолевая кислота и альфа- линоленовая (которые обычно группируют вместе под названием витамина F) заслуживает здесь особого внимания, поскольку их присутствие незаменимо для жизнедеятельности организма.
В последние несколько лет было показано, что эти жирные кислоты играют важнейшую роль в формировании мембран клеток головного мозга и в развитии нервной системы. Недостаток этих кислот приводит к нарушению функции головного мозга. Наиболее подвержены этой опасности молодые люди.
Итак напомним, что отсутствие необходимых жирных кислот может быть важным фактором в развитии наиболее серьезных заболеваний обмена веществ, поражающих население индустриального общества. Именно их отсутствие связывают с распространением заболеваний системы иммунной защиты.
Плохие привычки в питании и сомнительная природа продуктов, которые имеются в нашем распоряжении, особенно если они очищены, возможно являются причиной этой недостаточности. Линолевая кислота, которая найдена в подсолнечном, кукурузном маслах и масле из виноградных косточек снижает риск сердечно- сосудистых заболеваний.
Недостаток линолевой кислоты может привести к замедлению роста и к изменениям в клетках кожи, слизистой железы, желез внутренней секреции и половых органов. Рекомендуемая суточная доза — 10 граммов. Она может быть получена при употреблении 20 граммов подсолнечного, кукурузного или соевого масла.
Альфа-линоленовая кислота,найденная в больших количествах в маслах из рапсовых семян, маслах из грецкого ореха и зерен пшеницы особенно важна для биохимии нервной системы. Недостаточность этой кислоты может привести к снижению способности к обучению, отклонениям в передаче нервных сигналов, увеличению опасности возникновения тромбоза, а также к снижению сопротивления алкоголю. Ее ежедневно рекомендуемая доза — 2 грамма. Это доза может быть получена при ежедневном употреблении 25 граммов рапсового масла.
Никакое масло в отдельности не может дать правильного баланса олеиновой, линолевой и альфа- линоленовой кислот. Поэтому, вам следует смешивать два или три следующих различных масла как приправу к салату: оливковое масло, подсолнечное масло, рапсовое масло.
г. Ежедневное потребление липидов (жиров)
Ежедневное потребление жиров во всех их формах не должно составлять более 30% потребления пищевых продуктов. Во Франции в настоящее время оно составляет 45% (две трети которых состоят из насыщенных жиров).
Идеальным считается суточное потребление 25% насыщенных жиров (мясо, продукты свинины, масло, все молочные продукты), 50% мононенасыщенных жирных кислот (гусиный жир, оливковое масло) и 25% полиненасыщенных жирных кислот (рыба, подсолнечное масло, рапсовое масло, кукурузное масло и т.д
5. Строение и функции простых липидов – жира и воска.
Простые липиды в большинстве представлены сложными эфирами высших жирных кислот и трехатомного спирта глицерина — триглицеридами. Жирные кислоты имеют: 1) одинаковую для всех кислот группировку — карбоксильную группу (–СООН) и 2) радикал, которым они отличаются друг от друга. Радикал представляет собой цепочку из различного количества (от 14 до 22) группировок –СН2–. Иногда радикал жирной кислоты содержит одну или несколько двойных связей (–СН=СН–), такую жирную кислоту называют ненасыщенной. Если жирная кислота не имеет двойных связей, ее называют насыщенной. При образовании триглицерида каждая из трех гидроксильных групп глицерина вступает в реакцию конденсации с жирной кислотой с образованием трех сложноэфирных связей.
Среди веществ растительного и животного происхождения очень распространены соединения, относящиеся к классу сложных эфиров, т. е. вещества, структуру которых в наиболее общем виде можно представить как
R' — (CO) — O — R''
где R' и R" - углеводородные радикалы, которые могут иметь самое разное строение и длину.
Природные соединения такого состава называются липидами. К липидам относятся, во-первых, хорошо известные жиры и масла, относящиеся к сложным эфирам, образованным трехатомным спиртом - глицерином и высшими карбоновыми кислотами. К липидам относят и воски, очень схожие с жирами как химически, так я по ряду физических свойств.
Воски — соединения, образованные высшими карбоновыми кислотами и высокомолекулярными одноатомными спиртами различного строения.
Состав
В состав воска входят около 300 различных веществ, среди которых преобладают сложные эфиры, углеводороды, свободные жирные кислоты, ароматические вещества, вода, красящие, минеральные и другие вещества.
Главной составной частью воска являются сложные эфиры (до 75%), образованные пальмитиновой, неоцеротиновой и мелиссиновой кислотами, цериловым и мелиссиновым спиртами. Кроме того, в воске содержатся церотиновая, монтаниновая и олеиновая кислоты и неоцериловый и монтаниновый спирты. Церотиновая и мелиссиновая кислоты — наиболее активная часть воска; они могут вступать в реакцию с большинством металлов, а также со щелочами.
Физические свойства
За физические свойства воска отвечают такие параметры как температура плавления и застывания, коэффициент твердости, плотность, степенью вязкости.
Температура плавления (переход от твердого состояния в жидкое) находится в пределах от 61 до 65 градусов. Температура застывания (переход из жидкого состояния в твердое) колеблется в пределах 61 – 63 градусов. Следует отметить, что застывает воск при температуре несколько более низкой (на 0,1- 2 градуса), чем плавится. Если опустить термометр в воск во время, то в начале термометр показывает постепенно снижение температуры. Далее, достигнув примерно 61 – 63 градусов, она некоторое время остается неизменной, после чего происходит затвердевание воска. Остановка на короткий промежуток времени понижения температуры вызывают скрытой теплотой плавления. Именно эту температуру и принято считать температурой плавления воска.
Плотность воска показывает массу одного кубического сантиметра воска. Во время повышения температуры воска на один градус плотность воска уменьшается на 0,0008. Это может говорить о том, что плотность воска зависит от температуры. При нормальных условиях плотность чистого воска колеблется от 0,96 до 0,98. Твердый воск способен плавать на поверхности воды, но в расплавленном воске он тонет. Знание плотности воска важно при определении фальсификации натурального воска.
За твердость воска отвечает коэффициент твердости - время прохождения иглы с поперечным сечением 1,5 квадратного миллиметра под действием груза весом 1 килограмм на глубину в 1 миллиметр. Определение ведут всегда при температуре 20. Для светлого, без каких-либо примесей воска коэффициент твердости составляет 9 – 13. У извлеченного из старых сотов воска коэффициент твердости составляет примерно 4 – 6 Твердость воска очень важное физическое свойство: при производстве вощины: чем тверже воск, тем прочнее вощина. Твердость воска зависит от его качества. При изготовлении вощины рекомендуется применять воск с высоким коэффициентом твердости.
За вязкость воска отвечает сила сцеплением между его внутренними частицами. С помощью ее находят консистенцию и чистоту расплавленного воска. Что бы определить вязкости расплавленного воска, его при постоянной температуре пропускают через маленькие отверстия.
Химический состав. По химическому составу пчелиный воск представляет смесь различных веществ, количество которых колеблется в зависимости от происхождения воска, пищи пчел и условий добывания его из воскового сырья.
Из химических элементов в состав воска входят: углерод- около 80%, водород - около 13%, кислород -7%. Эти элементы объединены в 15 различных веществ. Главной составной частью воска являются сложные эфиры (70,4- 74,7%), образованные предельными одноосновными кислотами -(пальмитиновой, неоцеротиновой и мелиссиновой) и предельными одноосновными спиртами (цериловым и мелиссиновым). В меньших количествах в воске содержатся церотиновая, монтаниновая и олеиновая кислоты, а также спирты-неоцериловый и монтаниновый. Церотиновая и мелиссиновая кислоты представляют наиболее активную часть воска, они могут вступать в реакцию с большинством металлов и со щелочами.
Кроме того, пчелиный воск содержит предельные углеводороды (12,5-15,5%) и около 15% свободных кислот. В нем также есть вещества, обусловливающие его цвет и запах, а также минеральные соли.
Цвет воска зависит от красящих веществ, переходящих в него из прополиса и некоторых сортов пыльцы. Кроме того, на цвет воска влияет способ переработки воскового сырья. Ароматические вещества переходят в воск главным образом из меда, т. е. происходят из тех растений, с которых пчелы собирали нектар.
Растворителями воска являются вещества, не соединяющиеся с водой: бензин, скипидар, толуол, хлороформ и др. В спирте растворяется лишь очень небольшая часть веществ, входящих в состав воска. В воде и глицерине воск не растворяется, однако при определенных условиях он образует с водой эмульсию. Для образования эмульсии воска с водой обязательно присутствие третьего вещества, называемого эмульгатором. Такими эмульгаторами могут быть щелочи, соли жесткой воды, декстринообразные соединения меда, перга и другие вещества.
Свободные жирные кислоты воска растворяют многие металлы, образуя в большинстве случаев окрашенные соли. От соприкосновения с железом воск приобретает бурую окраску, а при сильном перегреве может даже совсем почернеть. Чугун действует на воск немного слабее, чем железо. Медь окрашивает воск в зеленый цвет. Цинк растворяется в воске, отчего он приобретает серовато-грязный цвет.
В фосфолипидах одна из крайних цепей высших карбоновых кислот триацилглицерола замещена на группу, содержащую фосфат. Фосфолипиды имеют полярные головки и неполярные хвосты. Группы, образующие полярную головку, гидрофильны, а неполярные хвостовые группы гидрофобны. Двойственная природа этих липидов обусловливает их ключевую роль в организации биологических мембран.
Еще одну группу липидов составляют стероиды (стеролы). Эти вещества построены на основе спирта холестерола. Стеролы плохо растворимы в воде и не содержат высших карбоновых кислот. К ним относятся желчные кислоты, холестерол, половые гормоны, витамин D и др.
К липидам также относятся терпены (ростовые вещества растений - гиббереллины; каротиноиды - фотосинтетичские пигменты; эфирные масла растений, а также воска).
В миелиновых оболочках аксонов нервных клеток липиды являются изоляторами при проведении нервных импульсов. Воск используется пчелами в строительстве сот.
И витаминов (ОПК-6)
План:
1. Строение, свойства и классификация аминокислот.
2. Строение, свойства и функции нуклеотидов.
3. Общая схема строения полипептидов.
4. Первичная, вторичная, третичная и четвертичная структура белков и её биологическое значение.
5. Содержание и состав белков в продуктах растительного и животного происхождения.
6. Строение, свойства и классификация витаминов.
Среди органических веществ клетки белки занимают первое место, как по количеству, так и по значению. У животных на них приходится около 50% сухой массы клетки. В организме человека встречается около 5 млн. типов белковых молекул, отличающихся не только друг от друга, но и от белков других организмов. Несмотря на такое разнообразие и сложность строения, белки построены всего из 20 различных аминокислот.
Содержание и состав белков в продуктах растительного и животного происхождения.
Белки бывают животного и растительного происхождения. Животные белки (мясо, рыба, птица, молочные продукты, яйца), а также некоторые растительные (соя, фасоль, горох), являются полноценными и должны составлять 60% от общего количества белков в сутки. Большинство растительных белков (например, цельные злаки) являются неполноценными, их доля равна 40%. Ниже представлена таблица продуктов животного и растительного происхождения, богатых белками.
Таблица 1. Содержание белков в продуктах животного происхождения
| Продукты | Белки, г/100 г (продукта) |
Мясо, субпродукты, колбасы
Птица, яйца
Рыба, морепродукты
Молочные продукты
Таблица 2. Содержание белков в продуктах растительного происхождения
| Продукты | Белки, г/100 г (продукта) |
|
Бобовые, орехи,семечки | |
| Соя | 34,9 |
| Чечевица | 24,8 |
| Горох | 23 |
| Фасоль | 22,3 |
| Семечки тыквы | 30,2 |
| Арахис | 26,3 |
| Семечки подсолнечника | 20,8 |
| Миндаль | 18,6 |
| Кешью | 18,2 |
| Фисташки | 20,3 |
| Фундук | 16,1 |
| Бразильский орех | 14,3 |
| Грецкий орех | 13,6 |
|
Овощи, грибы, сухофрукты | |
| Картофель | 2 |
| Свекла | 1,5-2,5 |
| Капуста брюссельская | 4,8 |
| Каруста кольраби | 2,8 |
| Капуста цветная | 2,5 |
| Капуста белокачанная | 1,8 |
| Чеснок | 6,5 |
| Шпинат | 2,9 |
| Редька | 1,9 |
| Репа | 1,5 |
| Морковь | 1,4 |
| Перец (красный сладкий) | 1,3 |
| Перец (зеленый сладкий) | 1,3 |
| Баклажаны | 1,2 |
| Грибы (белые) | 3,7 |
| Шампиньоны | 4,3 |
| Курага | 5,3 |
| Финики | 2,5 |
| Чернослив | 2,3 |
| Изюм | 1,9 |
Стоит отметить, что за один прием пищи наш организм способен усвоить не более 30 г белка. Кроме того степень усвоения различных белков весьма отличается. Она зависит как от типа белка (животный или растительный), так и от способа обработки пищи, общего состояния организма. Животные белки усваиваются примерно на 70-90%. Растительные - на 40-70%. Вот несколько примеров:
- Белки куриного яйца - степень усвоения 92-100%;
- Белки сквашенного молока - до 90%;
- Белки свежего молока - 83%;
- Белки говядины - 76%;
- Белки творога - 75%;
- Белки овсяной крупы - 66%;
- Белки изделий из пшеничной муки - 52%.
Функции (ОПК-6)
План:
1. Строение и общие свойства ферментов.
2. Основы современной классификации ферментов.
3. Принципы регуляции ферментативных реакций.
Биосинтез гликогена.
Гликоген – биополимер, состоящий из остатков глюкозы, он является компонентом всех тканей животных и человека (см. рисунок 15.1). Этот полисахарид служит основным источником энергии и резервом углеводов в организме.
Содержание гликогена в различных органах зависит от физиологического состояния организма. Наиболее высокое содержание гликогена обнаруживается в печени (от 2 до 6% от массы органа). Хотя концентрация гликогена в мышцах значительно ниже (от 0,5 до 1,5%), однако в норме его количество в мышцах составляет 2/3 от общего его содержания в организме.
Гликоген отличается значительным разнообразием по структуре и по относительной молекулярной массе. Молекулы его ветвисты. Глюкозные остатки образуют цепи, в которых они связаны между собой α-1,4-гликозидными связями. Разветвления образуются при помощи α-1,6-гликозидных связей.
Большая часть глюкозы, поступающей в организм с пищей, превращается в клетках печени в гликоген. Это связано с тем, что накопление легко растворимой глюкозы в клетках привело бы к резкому увеличению осмотического давления и разрушению клеточной мембраны.
Молекула глюкозы фосфорилируется при участии фермента глюкокиназы (в гепатоцитах) или гексокиназы (в других клетках) с образованием глюкозо-6-фосфата. Этот метаболит под влиянием фосфоглюкомутазы превращается в глюкозо-1-фосфат (рисунок 16.7, реакция 1).
Глюкозо-1-фосфат взаимодействует с уридинтрифосфатом (УТФ) в присутствии фермента УДФ-глюкозо-пирофосфорилазы с образованием УДФ-глюкозы – ключевого метаболита углеводного обмена (рисунок 7, реакция 2).
Перенос глюкозного остатка с УДФ-глюкозы на затравочную полисахаридную цепь осуществляет гликогенсинтаза – регуляторный фермент данного метаболического пути (рисунок 7, реакция 3).
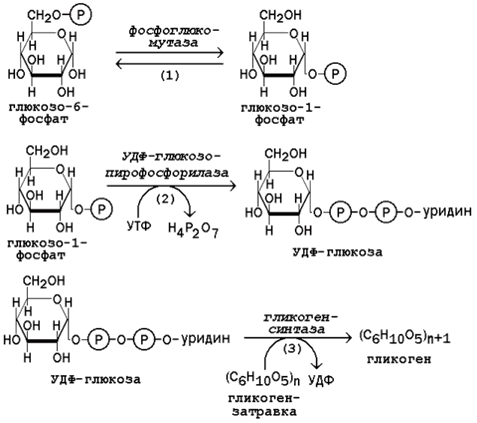
Рисунок 7. Реакции биосинтеза гликогена.
Гликогенсинтаза существует в двух формах: гликогенсинтаза b (неактивная) и гликогенсинтаза а (активная). Превращение формы а в форму b катализируется цАМФ-зависимой протеинкиназой, обратный процесс – протеинфосфатазой.
Образование α-1,6-гликозидных связей в молекуле гликогена катализирует ветвящий фермент или трансглюкозидаза (рисунок 8) .
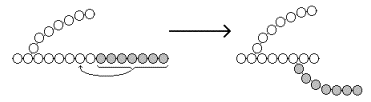
Рисунок 8. Образование α-1,6-гликозидных связей в молекуле гликогена.
| Мобилизация гликогена и гликогенолиз. |
Мобилизация, или распад гликогена – процесс превращения. Первую реакцию внутриклеточного расщепления гликогена катализирует фермент фосфорилаза гликогена (рисунок 9, реакция 1). Простетической группой его является пиридоксальфосфат (производное витамина В6). Фосфорилаза гликогена – регуляторный фермент, он существует в двух формах: фосфорилаза а (активная) и фосфорилаза b (неактивная).
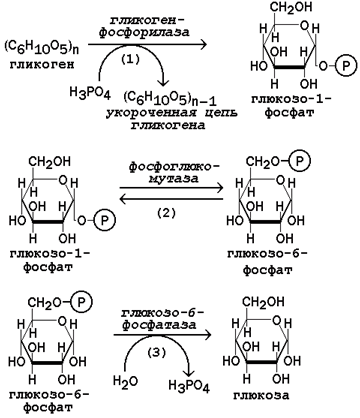 Рисунок 9. Реакции мобилизации гликогена.
Переход фосфорилазы b в фосфорилазу акатализирует киназа фосфорилазы, которая фосфорилирует неактивный фермент.
Превращение фосфорилазы а в фосфорилазу b катализирует протеинфосфатаза, которая осуществляет дефосфорилирование активного фермента. Следует отметить, что ускорение процессов мобилизации гликогена происходит одновременно с торможением его биосинтеза, и наоборот. Активацию фосфорилазы гликогена вызывают гормоны адреналин и глюкагон.
Фосфорилаза расщепляет в молекуле гликогена лишь α-1,4-гликозидные связи. Расщепление α-1,6-гликозидных связей осуществляется амило-1,6-гликозидазой гидролитически с образованием свободной глюкозы. Глюкозо-1-фосфат, образующийся в фосфорилазной реакции, при участии уже известного Вам фермента фосфоглюкомутазы переходит в глюкозо-6-фосфат (рисунок 9, реакция 2).
Дальнейшая судьба глюкозо-6-фосфата, образовавшегося при распаде гликогена, в разных тканях различна.
Как уже упоминалось, в печени и почках имеется фермент глюкозо-6-фосфатаза, отщепляющая фосфат от глюкозо-6-фосфата (рисунок 16.9, реакция 3). Образовавшаяся глюкоза диффундирует в кровь, откуда поглощается клетками организма и служит для них источником энергии.
В мышечной ткани фермент глюкозо-6-фосфатаза отсутствует. Поэтому глюкозо-6-фосфат, образовавшийся при распаде гликогена, подвергается дальнейшему превращению по гликолитическому пути с образованием лактата (этот процесс называется гликогенолизом). Образующийся в реакциях субстратного фосфорилирования АТФ используется в процессах мышечного сокращения. Таким образом, гликоген мышц является резервом энергии только для мышечной ткани.
Рисунок 9. Реакции мобилизации гликогена.
Переход фосфорилазы b в фосфорилазу акатализирует киназа фосфорилазы, которая фосфорилирует неактивный фермент.
Превращение фосфорилазы а в фосфорилазу b катализирует протеинфосфатаза, которая осуществляет дефосфорилирование активного фермента. Следует отметить, что ускорение процессов мобилизации гликогена происходит одновременно с торможением его биосинтеза, и наоборот. Активацию фосфорилазы гликогена вызывают гормоны адреналин и глюкагон.
Фосфорилаза расщепляет в молекуле гликогена лишь α-1,4-гликозидные связи. Расщепление α-1,6-гликозидных связей осуществляется амило-1,6-гликозидазой гидролитически с образованием свободной глюкозы. Глюкозо-1-фосфат, образующийся в фосфорилазной реакции, при участии уже известного Вам фермента фосфоглюкомутазы переходит в глюкозо-6-фосфат (рисунок 9, реакция 2).
Дальнейшая судьба глюкозо-6-фосфата, образовавшегося при распаде гликогена, в разных тканях различна.
Как уже упоминалось, в печени и почках имеется фермент глюкозо-6-фосфатаза, отщепляющая фосфат от глюкозо-6-фосфата (рисунок 16.9, реакция 3). Образовавшаяся глюкоза диффундирует в кровь, откуда поглощается клетками организма и служит для них источником энергии.
В мышечной ткани фермент глюкозо-6-фосфатаза отсутствует. Поэтому глюкозо-6-фосфат, образовавшийся при распаде гликогена, подвергается дальнейшему превращению по гликолитическому пути с образованием лактата (этот процесс называется гликогенолизом). Образующийся в реакциях субстратного фосфорилирования АТФ используется в процессах мышечного сокращения. Таким образом, гликоген мышц является резервом энергии только для мышечной ткани.
|
3. Механизмы образования глицерина, насыщенных и ненасыщенных жирных кислот.
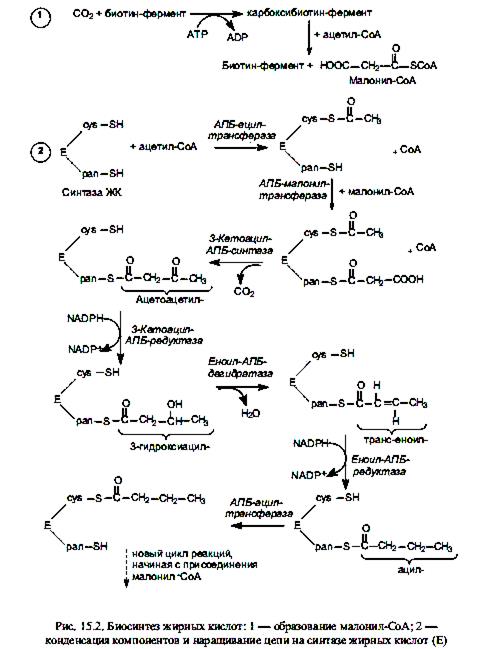
Биосинтез жирных кислот осуществляется в цитоплазме эукариотических клеток, у животных — в основном в клетках печени, жировой ткани, почек, легких. Непосредственным предшественником их синтеза является малонил-СоА, который образуется из ацетил-СоА. В свою очередь, основное количество ацетил-СоА образуется в матриксе митохондрий при окислительном декарбоксилировании пирувата, в процессе b-окисления жирных кислот, при расщеплении углеродных скелетов аминокислот. Будучи заряженным соединением, ацетил-СоА не может преодолеть мембранный барьер митохондрий, и для транспорта ацетильных групп в цитоплазму существует специальный «челночный механизм». С помощью этого механизма ацетильные компоненты попадают в цитоплазму в составе цитрата, который расщепляется, образуя ацетил-СоА (при участии АТР и СоА) и оксалоацетат. Последний восстанавливается в малат и возвращается в матрикс митохондрий, где регенерируется цитрат.
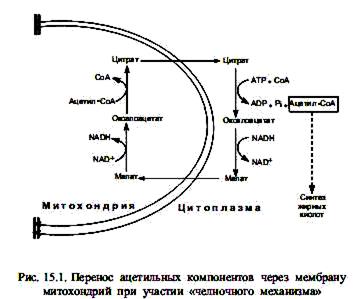
Оказавшись в цитоплазме, ацетил-СоА подвергается карбоксилированию с участием ключевого фермента биосинтеза жирных кислот—ацетил-СоА-карбоксилазы. В результате этой АТР-зависимой реакции формируется малонил-СоА. Простетической группой ацетил-СоА-карбоксилазы служит биотин. Он выполняет роль «подвижной руки», переносящей СО2 на молекулу ацетил-СоА. Реакция осуществляется в две стадии. Стадия образования малонил-СоА лимитирует скорость всего процесса синтеза жирных кислот, поскольку на уровне ацетил-СоА-карбоксилазы осуществляется регуляция данного пути: основным аллостерическим активатором фермента является цитрат. Этот метаболит образуется в митохондриях в большом количестве, когда там много ацетил-СоА, т. е. ЦТК перегружен «топливом», и его избыток должен запасаться в виде триацилглицеролов. В таком случае цитрат выходит в цитоплазму, выступая там одновременно в роли донора ацетил-СоА и активатора ацетил-СоА-карбоксилазы. Связывание цитрата с неактивными мономерами ацетил-СоА-карбоксилазы приводит к их соединению в нитевидный олигомер, который приобретает функциональную активность. Кроме этого, активность ацетил-СоА-карбоксилазы регулируется в ходе ковалентной модификации: понижается при фосфорилировании и повышается при дефосфорилировании.
Ненасыщенные жирные кислоты синтезируются из насыщенных с соответствующей длиной цепи. Образование двойной связи у аэробных организмов катализируется ферментом ацил-СоА-оксигеназой, и окисление происходит согласно реакции
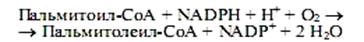
В этом процессе одновременно окисляются два разных субстрата: жирная кислота (возникает двойная связь) и NADPH, его катализируют монооксигеназы, называемые десатуразами. Известен и другой механизм образования двойных связей в молекулах жирных кислот, не требующий участия молекулярного кислорода. Например, в клетках E.coli синтез пальмитолеиновой кислоты начинается еще на синтазе жирных кислот со стадии образования двойной связи в составе С10-фрагмента. В этой реакции принимает участие особый фермент, присутствующий в клетках кишечной палочки,—b-оксидеканоилтиоэфирдегидратаза, который катализирует формирование цис-b,g-двойной связи, а не транс-a,b-, как это имеет место на стадии образования еноил-производного при синтезе насыщенных жирных кислот. Затем происходит удлинение ненасыщенного фрагмента до С16- и С18-производных.
У животных и растений введение в молекулу насыщенной жирной кислоты первой двойной связи осуществляется в цитозоле довольно легко. Образование дополнительных двойных связей у растений происходит в эндоплазматическом ретикулуме, а у животных не происходит вообще.
Поэтому полиненасыщенные жирные кислоты (линолевая, линоленовая) являются для животных, в частности млекопитающих, незаменимыми компонентами и должны поступать в организм с пищей (растительного происхождения). Эти жирные кислоты служат субстратами для построения других полиненасыщенных жирных кислот. Недостаток линолевой и леноленовой кислот в рационе животных приводит к торможению роста, поражению кожных покровов и почек, нарушению функции размножения.
Биосинтез липидов в клетках эукариот осуществляется на мембранах гладкого эндоплазматического ретикулума. Большинство ферментов, принимающих участие в этих процессах, ассоциированы с его мембранами и представляют собой липопротеины.
Основными предшественниками для синтеза нейтральных липидов (триацилглицеролов), а также полярных фосфо- и гликолипидов служат активированные жирные кислоты и глицерол-3-фосфат. Активация жирных кислот происходит в следующей реакции:
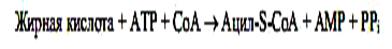
Глицерол-3-фосфат образуется либо при прямом фосфорилировании за счет АТР при участии глицеролкиназы, либо при восстановлении промежуточного продукта гликолиза — дигидроксиацетон-3-фосфата ферментом 3-глицерофосфатдегидрогеназой, использующей в качестве кофермента NADH.
Биосинтез неполярных липидов. Если биосинтез триацилглицеролов осуществляется de novo (из глицеролфосфата и жирных кислот), то на первой стадии происходит последовательное ацилирование двух свободных гидроксильных групп молекулы глицерол-3-фосфата (рис. 15.3). При этом в реакции этерификации СоА-эфиром жирной кислоты по первому атому углерода глицерол-3-фосфата вначале формируется моноацилглицерол-3-фосфат (лизофосфатид), а затем — диацилглицерол-3-фосфат (фосфатидат).
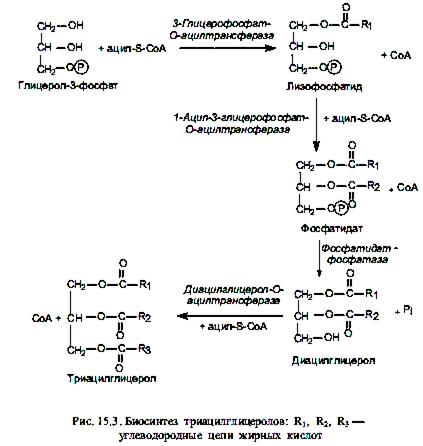
На следующей стадии происходит гидролитическое отщепление фосфатной группы от молекулы фосфатидата и образуется 1, 2-диацилглицерол, который взаимодействует с третьим СоА-производным жирной кислоты, в результате чего формируется триацилглицерол (триглицерид).
Кроме описанной схемы, синтез нейтральных липидов может осуществляться с участием в качестве предшественников продуктов расщепления липидов, попадающих в организм с пищей. Эти процессы особенно интенсивны в слизистой кишечника животных. Нейтральные жиры расщепляются в пищеварительном тракте панкреатическими липазами до жирных кислот и 2-моноацилглицеролов, которые всасываются слизистой кишечника. В клетках слизистой оболочки происходит последовательное ацилирование 2-моноацилглицерола СоА-эфирами жирных кислот с образованием триацилглицеролов. Эти реакции катализируют особые ацилтрансферазы.
Триглицериды, как уже отмечалось, являются основными запасными веществами в клетках животных и некоторых других организмов. Особое значение они имеют для впадающих в спячку и мигрирующих на далекие расстояния животных. Например, верблюды запасают триацилглицеролы в горбу и используют их как источник воды, которая образуется при окислении. У полярных животных (тюленей, моржей и др.) триглицериды часто выполняют функцию теплоизолятора. Некоторые животные используют неполярные липиды для регулирования плавучести. Например, в спермацетовом мешке кашалотов находится несколько тонн триацилглицеролов, содержащих в составе молекул ненасыщенные жирные кислоты. Плотность (консистенция) этих триглицеридов зависит от температуры среды: повышается при понижении температуры. Питаясь кальмарами, кашалоты заплывают на большие глубины, где температура воды ниже обычной. Это индуцирует кристаллизацию триглицеридов, увеличивается их плотность соответственно увеличению плотности морской воды на глубине, и животное, не прибегая к дополнительным усилиям, может долго оставаться на большой глубине.
Образование углеводов из продуктов глиоксилатного цикла.
Глиоксила́тный цикл , или глиоксила́тный шунт — анаболический путь, имеющийся у растений, бактерий, протистов и грибов, представляет собой видоизменённый цикл трикарбоновых кислот. Глиоксилатный цикл служит для превращения ацетил-СоА в сукцинат, который далее используется для синтеза углеводов. У микроорганизмов он обеспечивает утилизацию простых углеродных соединений в качестве источника углерода, когда более сложные источники, например, глюкоза, недоступны, а также может считаться одной из анаплеротических реакций цикла трикарбоновых кислот, восполняющей количества сукцината и малата. Считается, что у животных глиоксилатный цикл отсутствует (подтверждённое исключение составляют нематоды на ранних стадиях своего эмбриогенеза), однако в последние годы в некоторых тканях животных были обнаружены ключевые ферменты цикла — малатсинтаза и изоцитратлиаза.
Ферменты и реакции глиоксилатного цикла были открыты Хансом Корнбергом и Хансом Адольфом Кребсом.
У растений, некоторых беспозвоночных и некоторых микроорганизмов (в том числе бактерии Escherichia coli и дрожжей) ацетат может служить как источником энергии, так и предшественником фосфоенолпирувата, который далее используется для синтеза углеводов. У этих организмов ферменты глиоксилатного цикла катализируют превращение ацетата в сукцинат:
2 Ацетил-СоА + NAD+ + 2H2O → сукцинат + 2СоА + NADH + H+.
В ходе глиоксилатного цикла ацетил-СоА соединяется с оксалоацетатом с образованием цитрата, а цитрат превращается в изоцитрат, как и в цикле трикарбоновых кислот. Однако следующим шагом является не отщепление от изоцитрата водорода ферментом изоцитратдегидрогеназой, а расщепление изоцитрата на сукцинат и глиоксилат под дейстствием изоцитратлиазы. Глиоксилат после этого конденсируется со второй молекулой ацетил-СоА, образуя малат под действием малатсинтазы, а сукцинат покидает цикл. Малат далее окисляется в оксалоацетат, который может соединиться с новой молекулой ацетил-СоА и начать новый оборот цикла. В ходе каждого оборота глиоксилатного цикла используются две молекулы ацетил-СоА и образуется одна молекула сукцината, который в дальнейшем может использоваться в биосинтетических процессах. Сукцинат может быть преобразован через фумарат и малат в оксалоацетат, который далее может превратиться в фосфоенолпируват под действием фосфоенолпируваткарбоксикиназы, а фосфоенолпируват в ходе глюконеогенеза даёт глюкозу. У животных глиоксилатный цикл не происходит, и в связи с этим они неспособны получать глюкозу из липидов (ацетил-СоА образуется при β-окислении жирных кислот).
У растений ферменты глиоксилатного цикла находятся в мембраносвязанных органеллах — глиоксисомах, представляющих собой специализированные пероксисомы (названы по протекающему в них циклу). Ферменты, одновременно участвующие и в цикле трикарбоновых кислот, и в глиоксилатном цикле, имеют по два изофермента, один из которых локализуется в митохондриях, а другой — в глиоксисомах. Глиоксисомы не присутствуют постоянно во всех растительных тканях. Они образуются в богатых липидами семенах при их прорастании, ещё до того, как развивающееся растение сможет само образовывать глюкозу в ходе фотосинтеза. Кроме ферментов глиоксилатного цикла, в глиоксисомах находятся все ферменты, необходимые для деградации жирных кислот, запасённых к семенах. Ацетил-СоА, образующийся при β-окислении жирных кислот, превращается в сукцинат в ходе глиоксилатного цикла, и сукцинат транспортируется в митохондрии, где он вовлекается в цикл трикарбоновых кислот и превращается в малат. Цитозольный изофермент малатдегидрогеназы окисляет малат до оксалоацетата, который в свою очередь может превратиться в фосфоенолпируват. Последний вовлекается в глюконеогенез и даёт в конечном итоге глюкозу. Таким образом, прорастающее семя может переводить углерод, запасённый в липидах, в глюкозу.
У грибов ферменты глиоксилатного цикла локализуются в пероксисомах. У некоторых патогенных грибов глиоксилатный цикл может служить для совершенно других целей. Концентрация двух ключевых ферментов цикла — изоцитратлиазы и малатсинтазы — значительно возрастает при контакте гриба с человеком, которого этот гриб поражает. У некоторых видов таких грибов мутанты, лишённые изоцитратлиазы, имеют сниженную вирулентность по сравнению с диким типом, как показали исследования на мышах. Причины такой связи в настоящий момент исследуются, однако можно заключить, что глиоксилатный цикл необходим для поддержания вирулентности этих патогенов.
Согласно новейшим данным, у растений и грибов ферменты глиоксилатного цикла локализованы не только внутри глиоксисомы: некоторые из них находятся снаружи, поэтому протекание глиоксилатного цикла включает транспорт некоторых его промежуточных соединений через мембрану пероксисомы. Благодаря наличию глиоксилатного цикла некоторые микроорганизмы могут расти в среде, содержащей ацетат или жирные кислоты в качестве единственного источника углерода.
Принято считать, что животные неспособны к осуществлению глиоксилатного цикла, поскольку до недавнего времени не было никаких доказательств наличия у них двух ключевых ферментов цикла — изоцитратлиазы и малатсинтазы. Однако некоторые исследования показывают, что этот цикл имеется у некоторых, если не всех, позвоночных. В частности, некоторые исследования показали, что компоненты глиоксилатного цикла присутствуют в больших количествах в печени курицы. Эти и подобные им данные дают основания предполагать, что теоретически глиоксилатный цикл может осуществляться и более сложно организованными позвоночными. Другие исследования приводят доказательства того, что глиоксилатный цикл имеется у некоторых насекомых и морских беспозвоночных, а также нематод (в последнем случае наличие глиоксилатного цикла чётко подтверждено). Впрочем, другие исследования такие выводы опровергают. По поводу наличия цикла у млекопитающих также нет единого мнения: например, в одной работе указывается, что глиоксилатный цикл активен у медведей в зимней спячке, однако более поздние исследования оспаривают этот вывод]. С другой стороны, в геномах плацентарных млекопитающих не было идентифицировано ни одного гена, который кодировал бы изоцитратлиазу и малатсинтазу, хотя, по-видимому, малатсинтаза может быть активна у неплацентарных млекопитающих и других позвоночных. Более того, предполагается, что гены, кодирующие эти ферменты, у млекопитающих находятся в виде псевдогенов, то есть такие гены не совершенно отсутствуют у млекопитающих, а лишь находятся в «выключенном» состоянии. Возможно, в регуляцию глиоксилатного цикла у позвоночных вовлечён витамин D.
Впрочем, в отсутствии глиоксилатного цикла в клетках млекопитающих есть и свои выгоды. Наличие глиоксилатного цикла у некоторых патогенных микроорганизмов и отсутствие его у людей даёт возможности для создания антибиотиков, блокирующих это цикл: они были бы губительны для зависимых от глиоксилатного цикла микроорганизмов и при этом безвредны для людей.
В прорастающих семенах ферментативные превращения дикарбоновых и трикарбоновых кислот происходят в трёх внутриклеточных компартментах: митохондриях, глиоксисомах и цитозоле. Между этими компартментами существует постоянный обмен метаболитами.
Углеродный скелет оксалоацетата из цикла трикарбоновых кислот (протекает в митохондриях) переносится в глиоксисомы в форме аспартата. Там аспартат превращается в оксалоацетат, который конденсируется с ацетил-СоА, образовавшимся при окислении жирных кислот. Образующийся при этом цитрат под действием аконитазы переводится в изоцитрат, который далее распадается на глиоксилат и сукцинат под действием изоцитратлиазы. Сукцинат возвращается в митохондрию, где он вовлекается в цикл трикарбоновых кислот и превращается в малат, который переносится в цитоплазму и под действием цитозольной малатдегидрогеназы окисляется в оксалоацетат. В ходе глюконеогенеза оксалоацетат превращается в гексозы и сахарозу, которые могут доставляться в растущие корни и побег. Таким образом, во всех этих превращениях задействованы 4 различных биохимических процесса: окисление жирных кислот до ацетил-СоА (в глиоксисомах), глиоксилатный цикл (в глиоксисомах), цикл трикарбоновых кислот (в митохондрии) и глюконеогенез (в цитозоле).
Наличие общих метаболитов у этих процессов требует их координированной регуляции. Важнейшим из этих соединений является изоцитрат, служащий «точкой расхождения» глиоксилатного цикла и цикла трикарбоновых кислот. Изоцитратдегидрогеназа регулируется при помощи ковалентных модификаций: специфическая протеинкиназа фосфорилирует её и тем самым инактивирует. Эта инактивация приводит к тому, что изоцитрат не может далее участвовать в цикле трикарбоновых кислот и направляется в глиоксилатный цикл, где он становится предшественником для синтеза глюкозы. Фосфопротеинфосфатаза удаляет фосфорильную группу с изоцитратдегидрогеназы, вновь активируя фермент, и большая часть изоцитрата направляется в цикл трикарбоновых кислот, где из него извлекается энергия. Регуляторные ферментативные активности протеинкиназы и фосфопротеинфосфатазы осуществляются одним полипептидом.
Некоторые бактерии, в том числе E. coli, имеют в цитозоле полный набор ферментов гликосилатного цикла и цикла трикарбоновых кислот и поэтому могут расти на ацетате как единственном источнике углерода и энергии. Фосфопротеинфосфатаза, активирующаяся изоцитратдегидрогеназу, стимулируется промежуточными соединениями цикла трикарбоновых кислот и гликолиза, которые служат индикаторами того, что клетка в достаточной мере обеспечена энергией. Те же метаболиты подавляют протеинкиназную активность этого регуляторного полипептида. Поэтому накопление промежуточных продуктов центральных путей, обеспечивающих клетку энергией, свидетельствующее о нехватке энергии, активирует изоцитратдегидрогеназу. Когда концентрация этих веществ падает, свидетельствуя о достаточном снабжении исходными продуктами цикла трикарбоновых кислот, изоцитратдегидрогеназа инактивируется протеинкиназой.
Те же соединения гликолиза и цикла трикарбоновых кислот, которые активируют изоцитратдегидрогеназу, являются аллостерическими[en] ингибиторами изоцитратлиазы. Когда скорость энергетического обмена достаточно высока, чтобы поддерживать концентрацию промежуточных соединений глиоксилатного цикла и цикла трикарбоновых кислот низкой, изоцитратдегидрогеназа инактивирована, а ингибирование изоцитратлиазы прекращается, и изоцитрат вовлекается в глиоксилатный цикл, где он будет использован для биосинтеза углеводов, аминокислот и других клеточных компонен
Происхождения (ПК-22)
План:
1. Биохимическая характеристика органических кислот.
2. Влияние природно-климатических факторов, удобрений и других условий выращивания на накопление органических кислот в растительных продуктах. 3. Общая характеристика вторичных метаболитов.
Курс лекций
по дисциплине «Биохимия сельскохозяйственной продукции»
(для бакалавров технологического факультета по направлению: 35.03.07 -
Технология производства и переработки сельскохозяйственной
продукции)
Тверь, 2017
Составитель: к. с.-х. н., доцент кафедры агрохимии и земледелия
Шилова О.В.
Рецензент: к. б. н., доцент кафедры биологии животных, зоотехнии и основ
ветеринарии Тверской ГСХА Корецкая Е.А.
Шилова О.В. Курс лекций по дисциплине «Биохимия сельскохозяйственной продукции» (для бакалавров технологического факультета по направлению: 35.03.07 - Технология производства и переработки сельскохозяйственной
продукции) – Тверь, 2017. – 152 с.
Председатель методической комиссии Акимов А.А.
Введение
Цели дисциплины: формирование современных представлений, знаний и умений о превращениях веществ и энергии в живых организмах, химическом составе сельскохозяйственной продукции растительного и животного происхождения, биохимических процессах, происходящих в ней при хранении и переработке.
Задачи дисциплины: изучение строения и биологических функций важнейших органических веществ; механизмов ферментативных и биоэнергетических превращений в организмах; химического состава сельскохозяйственной продукции и биохимических процессов, происходящих в ней при хранении и переработке;
– оценка качества и технологических свойств сельскохозяйственной продукции по биохимическим показателям;
– применение знаний о химическом составе и биохимических процессах при обосновании технологий производства, хранения и переработки сельскохозяйственной продукции;
– ознакомление с современными методами и достижениями биохимической науки.
Модуль 1. Характеристика и биологические функции
Органических соединений
Дата: 2018-12-21, просмотров: 910.