Ген – элементарный, дискретный, материальный наследственный фактор, который определяет строение одной белковой полипептидной цепи. Он является единицей наследственности и передается от родителей к их потомкам. Ген представляет собой определенную последовательность нуклеотидов в нуклеотидной последовательности цепи ДНК. Гены расположены линейно, друг за другом. Размеры генов неодинаковы. Они зависят от размеров белка, который кодирует данный ген: если белок состоит из 200 остатков аминокислот, то в гене будет 600 пар нуклеотидов (по свойству генетического кода – триплетность, три нуклеотида кодируют одну аминокислоту), что соответствует относительной молекулярной массе около 420000. В хромосоме млекопитающих и человека средняя длина ДНК составляет около 2 см, что соответствует около 2,9х109 пар нуклеотидов. Всего же в геноме человека (в каждой клетке) около 6х10-12 г ДНК, что соответствует общей длине около 2 м и молекулярной массе около 3,7х1012. Учитывая, что средняя длина структурного гена составляет 500-1000 нуклеотидов, ДНК человека теоретически могла бы кодировать около 10 млн белков. Сколько же всего генов, то есть последовательностей, кодирующих белки, имеется в составе человеческой ДНК? Тридцать лет назад называли цифру 100 тысяч. К 2000 году специалисты по биоинформатике начали склоняться к мысли, что на самом деле нужны более осторожные оценки и что в геноме человека может оказаться 50-60 тысяч генов. Это означает, что кодирующая часть генома составляет всего 10-15% от всей ДНК. Следовательно, в нашем геноме имеется огромный «избыток» ДНК. В настоящее время показано, что значительная часть этой «избыточной» ДНК содержит гены, не являющиеся структурными, т.е. не кодирующие структуру определенных белков, а выполняющие целый ряд функций. Связанных с регуляцией активности тех или иных участков генома. Помимо этого, многие, в особенности регуляторные, гены многократно повторяются, т.е. геном содержит много (иногда десятки тысяч) копий одного и того же гена. Наконец, довольно большие участки ДНК разделяют гены между собой, кодируют начало и конец транскрипции и выполняют ряд других функций, еще не полностью выясненных. Кроме того, следует помнить, что у многоклеточных, в частности у человека и млекопитающих, в результате сложного процесса дифференцировки в ходе индивидуального развития образуется много разнообразных органов и тканей. Состоят эти органы и ткани из многочисленных клеток, специализированных для выполнения соответствующих функций. Видимо, именно поэтому большая часть генов многоклеточных организмов находится в неактивном состоянии. По результатам программы «Геном человека», опубликованным 13 февраля 2001 года, только 1% генома приходится на кодирующие экзоны, 24% - на некодирующие интроны и 75% - на межгенные промежутки. Избыточная ДНК характерна для всех эукариот. С помощью молекулярно-биологического метода изучения кинетики ренатурации денатурированной ДНК показали наличие в геноме эукариот следующих фракций: 1) уникальные последовательности, т.е. представленные в одном экземпляре; 2) промежуточные (среднечастотные) – повторяющиеся десятки и сотни раз; 3) высокочастотные повторы, число которых в геноме достигает 106 копий.
Дальнейшее развитие учения о строении гена показало, что:
1. Кроссинговер может происходить только между генами и, таким образом, ген – элементарная единица генетической рекомбинации. Однако доказано, что рекомбинации происходят и внутри гена. Наименьшая единица рекомбинации названа РЕКОНОМ.
2. Ранее считали ген единицей мутации. Однако было обнаружено, что изменения отдельных участков внутри сложного гена приводят к изменению его функции. Мельчайшая единица, способная к изменению, была названа МУТОНОМ.
3. Ген считали единицей функции. Многочисленные исследования показали, что функция гена может изменяться в зависимости от того, расположены ли два мутантных аллеля сложного гена в одной хромосоме, а их нормальные аллели в гомологичной (цисположение), или мутантные аллели расположены в двух гомологичных хромосомах (трансположение). Единицу функции предложено назвать ЦИСТРОНОМ.
Параллельная работа биохимиков и генетиков показала, что наименьшая величина рекона и мутона близка к величине одного или нескольких нуклеотидов, поскольку изменение даже одного основания в ДНК способно вызвать так называемую точковую мутацию. Цистрон же гомологичен участку ДНК, кодирующему синтез определенного полипептида, и содержит тысячу и более нуклеотидов.
Главное свойство гена – его экспрессия (работа). Следует отметить, что механизм экспрессии генов длительно время считался соответствующим только формуле, предложенной Дж. Бидлом и Э. Тейтемом в 1941 г. («один ген – один фермент») и в дальнейшем преобразованной в формулу «один ген – одна полипептидная цепь». Эта формула длительное время считалась центральной догмой молекулярной биологии, однако затем произошло её дополнение другими формулами, объясняющими механизмы экспрессии генов, и она перестала так называться. В 1965 г. У. Дрейер и Дж. Беннет предложили для объяснения механизма биосинтеза иммуноглобулинов формулу «два гена – одна полипептидная цепь». Оказалось, что в случае биосинтеза антител один ген отвечает за синтез вариабельной области иммуноглобулина, а другой ген – за синтез константной области иммуноглобулина. Спустя 30 лет Р. Дулиттл (1995) предложил еще одну формулу: «семь генов – одна полипептидная цепь». Результаты его исследований показали, что белковый домен – синтетаза жирных кислот – имеет только одну полипептидную цепь, но содержит в ней все необходимое для катализа семи реакций, значит, налицо общий результат действия семи разных генов. В результате выполнения международной программы «Геном человека» право на существование теперь имеют и такие формулы: «один ген – две, три и более полипептидные цепи», а также «разные участки одного гена – разные полипептидные цепи». Например, мутации в разных частях онкогена RET обуславливают развитие четырех разных наследственных болезней.
Установлено, что в клетках синтез одних ферментов происходит непрерывно, а других только в присутствие определенных индукторов. На основании своих исследований Ф. Жакоб и Ж. Моно в 1961 году предложили схему регуляции синтеза белков (ферментов) у микроорганизмов и она получила название гипотезы оперона.
По этой схеме гены функционально неодинаковы: одни из них (структурные) содержат информацию о расположении аминокислот в белке, другие выполняют регуляторную функцию (регуляторные гены). Структурные гены (цистроны) обычно расположены рядом и образуют один блок – оперон. Кроме того, в оперон входят промотор – место первичного прикрепления РНК-полимеразы и регуляторный участок – ген-оператор. Оператор включает и выключает структурные гены, это функциональный ген. Оператор, в свою очередь кодируется геном регулятором. Инициатор – место начала считывания генетической информации. Структурные гены – определяют синтез белков-ферментов, обеспечивающие цепь последовательных биохимических реакций. Помимо оперона, в том же или другом месте генома имеется особый ген – регулятор. Этот ген кодирует образование особого белка – репрессора, соединяющегося с оператором и подавляющего функцию всего оперона.
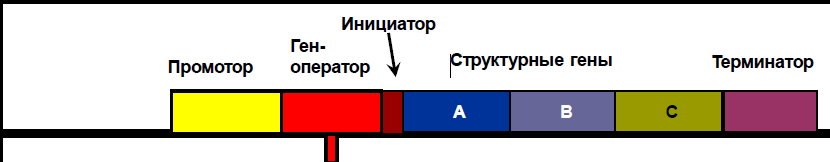
Регуляция по типу индукции. Вещество, поступившее в клетку – лактоза является индуктором, оно соединяется с белком репрессором лактозного оперона, переводит его в неактивную форму (инактивирует его), вследствие чего последний теряет способность подавлять функцию оперона. Оперон включается и начинается транскрипция со структурных генов, синтезируются ферменты, расщепляющие лактозу. Как только лактоза в среде будет расщеплена, репрессор освобождается, переходит в активную форму, блокирует оператор и работа структурных генов прекращается.
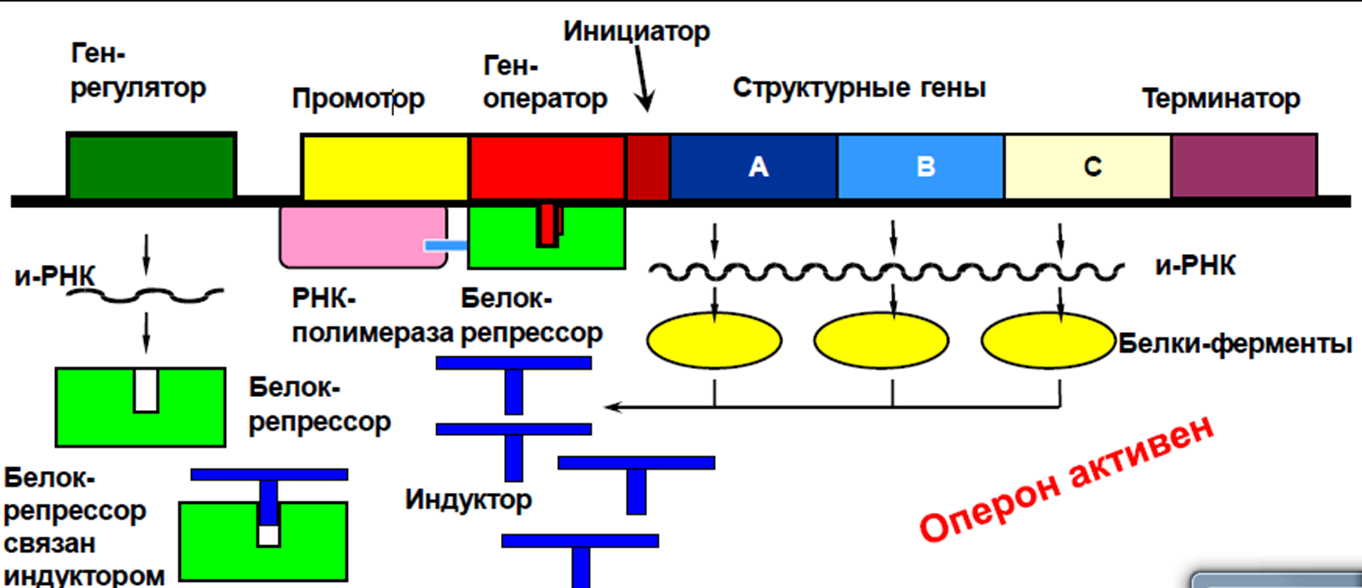
Регуляция по типу репрессии. Если в клетке в результате какой-то цепи реакций образуется конечный продукт в количестве большем, чем это необходимо, что может привести к нарушению обмена, то конечное вещество вступает в реакцию с белком репрессором, переводит его в активное состояние, происходит присоединение репрессора к гену – оператору, тем самым выключается вся система, и синтез ферментов прекращается.
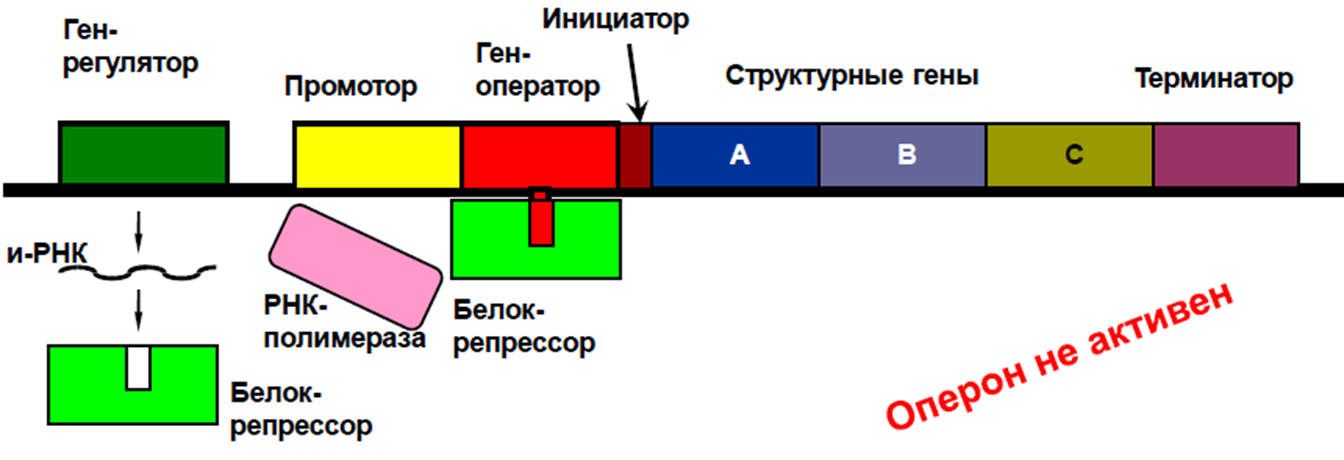
У высших организмов генетический контроль и регуляция биосинтеза белков много сложнее, чем у микроорганизмов. Прежде всего, хотя геном одинаков в разных клетках и тканях организма, в них синтезируются разные белки (миозин – в мышечных клетках, пепсин – в клетках слизистой желудка, гемоглобин – в эритробластах и т.д.). Это явление объясняется блокированием большей части генов в процессе дифференцировки, осуществляемым, по-видимому, при участии гистонов и других белков хроматина. Помимо этого, имеет место также регуляция. Связанная с индукцией ряда ферментов и других белков. Эта индукция у высших часто осуществляется на уровне всего организма при помощи гормонов, нервных импульсов и других механизмов.
Дата: 2018-11-18, просмотров: 1141.