В ходе исторического развития жизни на Земле первыми животными, у которых появилась нервная система, являются кишечнополостные. Это беспозвоночные двухслойные животные, типичным представителем является гидра пресноводная. Тело гидры представляет собой полый мешок, внутренняя полость является пищеварительной полостью. Наружный слой клеток называется эктодермой (в дословном переводе означает «наружная кожа»), а внутренний – энтодермой («внутренняя кожа»).
Нервные клетки гидры (рис. 14) расположены на границе между экто- и энтодермой. Они образуют простейшую нервную сеть диффузного типа. Каждая нервная клетка имеет длинные отростки и соединена с другими нервными клетками. Нервные клетки кишечнополостных являются изополярными, что означает отсутствие специализации у их отростков, а следовательно отростки проводят возбуждение в любую сторону и не образуют длинных проводящих путей. Контакты между нервными клетками в такой сети разнообразны:
анастомозы – плазматические контакты, обеспечивающие непрерывность сети
 щелевидные контакты – подобны синапсам. Бывают двух видов:
щелевидные контакты – подобны синапсам. Бывают двух видов:
o симметричные синапсы – содержат синаптические пузырьки по обе стороны контакта
o несимметричные синапсы – имеют везикулы только с одной стороны щели.
 Диффузный тип нервной системы характеризуется следующими признаками:
Диффузный тип нервной системы характеризуется следующими признаками:
нервные клетки равномерно распределены в теле животного. У кишечнополостных имеется два неоформленных скопления нервных клеток – в районе подошвы и ротового отверстия
проведение возбуждения во всех направлениях. Отсутствие специализированных отростков (дендритов и нейритов) связано с отсутствием специализированной рецепции. У гидры есть отдельные рецепторные клетки, но они не способны четко дифференцировать разные раздражители. Отсюда и отсутствие четко дифференцированной ответной реакции. Кишечнополостные способны избегать неблагоприятных факторов среды, не дифференцируя эти факторы (рис. 15).
волну распространяющегося возбуждения сопровождает волна мышечного сокращения. Вся дальнейшая эволюция нервной системы будет связана с эволюцией рецепторных и двигательных систем.
Вообще, для беспозвоночных животных характерно наличие нескольких источников происхождения нервных клеток. Для них возможно одновременное и независимое развитие изополярных нейронов из трех зародышевых листков (впрочем, происхождение нейронов из мезодермы до сих пор оспаривается, однако, есть работы, в которых сообщается о развитии нервных элементов из мезодермы у ряда примитивных беспозвоночных). Считается, что такой разнообразный нейрогенез является причиной многочисленности медиаторов нервных систем беспозвоночных.
Диффузный тип нервной системы характерен и для трехслойных животных – плоских червей. Однако в связи с более сложным строением тела - появлением третьего зародышевого листка (мезодермы – «промежуточной кожи»), двусторонней симметрии, примитивных органов чувств – 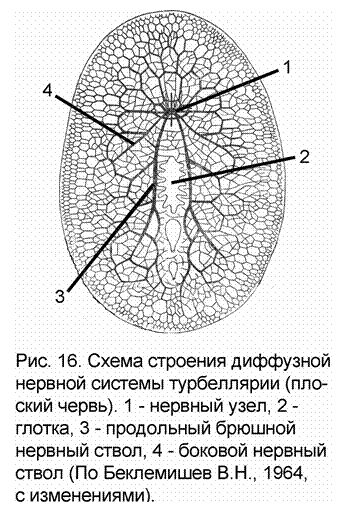 статоцистов (аналог органа равновесия), «глазков», обонятельных ямок (т.е. эволюцией рецепторного аппарата), - диффузная сеть усложняется. Из нее обособляются несколько продольных стволов, расположенных вдоль тела (рис. 16). В переднем конце животного эти стволы соединяются поперечными перемычками. Такая решетка нервных структур получила название ортогона. Стволы ортогона принципиально отличаются от нервных стволов (нервов) тем, что первые на всем своем протяжении содержат как нейроны, так и их отростки, а вторые состоят исключительно из отростков, а нейроны объединяются в ганглии.
статоцистов (аналог органа равновесия), «глазков», обонятельных ямок (т.е. эволюцией рецепторного аппарата), - диффузная сеть усложняется. Из нее обособляются несколько продольных стволов, расположенных вдоль тела (рис. 16). В переднем конце животного эти стволы соединяются поперечными перемычками. Такая решетка нервных структур получила название ортогона. Стволы ортогона принципиально отличаются от нервных стволов (нервов) тем, что первые на всем своем протяжении содержат как нейроны, так и их отростки, а вторые состоят исключительно из отростков, а нейроны объединяются в ганглии.
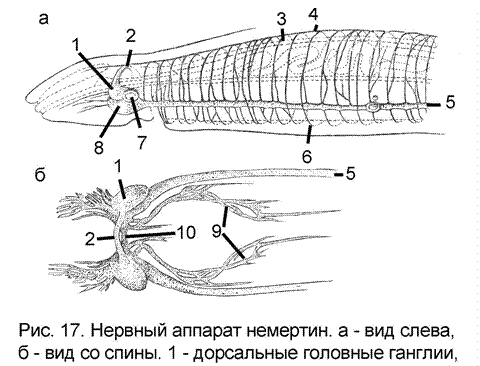
Общее направление эволюции нервного аппарата у низших червей – уменьшение числа нервных стволов и комиссур, уход нервного комплекса вглубь тела, возникновение церебрального (головного) ганглия (связано с развитием органов чувств, в частности статоциста, органов обоняния) – привело к внешнему архитектоническому упрощению нервного аппарата. Все перечисленное максимально выражено у немертин (сколециды - низшие черви), в мозгу которых появляются скопления ассоциативных клеток наподобие высших ассоциативных центров членистых животных (рис.17).
Развитие диффузной нервной системы в систему ортогонального типа определяет следующие направления эволюции нервного аппарата:
Централизация нервной системы.
Интеграция функций организма – интегрирующая роль нервного аппарата возрастает по мере увеличения степени его собственной централизации.
 |
5.2. Ганглионарный тип нервной системы
Появление этого типа нервной системы тесно связано с другим эволюционным нововведением – появлением сегментированных животных – кольчатых червей. Тело этих животных состоит из большого числа повторяющихся сегментов, или метамеров. В каждом сегменте имеется ганглий – парное скопление нервных клеток. Именно ганглии становятся основной анатомической структурой у большого количества таксономических групп животных. Кроме упомянутых кольчатых червей ганглионарный тип нервной системы типичен для двустворчатых, брюхоногих и головоногих моллюсков (рис. 18). У последних органы чувств, в частности глаза, достаточно хорошо развиты (рис. 19). Ганглионарный тип строения нервной системы типичен и для членистоногих.
Ганглии представляют собой скопления нервных клеток, окруженных соединительнотканной капсулой (рис. 18,в). Типичным для ганглиев является корковое строение: тела нейронов располагаются непосредственно под капсулой, направляя свои отростки внутрь ганглия. Центральная часть ганглия, состоящая из нервных отростков и глиальных элементов, называется нейропилем. Далее отростки нейронов выходят за пределы ганглия и формируют нервы и нервные стволы: комиссуры и коннективы. Комиссурами называют нервные стволы, объединяющие парные ганглии сегментов (если животное метамерно, например, кольчатые черви), или одноименные ганглии животных в значительной степени утративших метамерность (например, моллюски). Коннективы соединяют в цепочки ганглии соседних сегментов у метамерных животных или разноименные ганглии у неметамерных.
 В ходе эволюции ганглионарного типа нервной системы отмечаются следующие основные тенденции:
В ходе эволюции ганглионарного типа нервной системы отмечаются следующие основные тенденции:
дальнейшая централизация и интеграция нервной системы. Она проявляется в
o укорочении коннктив и комиссур
o слиянии ганглиев (одноименных и разноименных). Ганглии, образовавшиеся путем слияния имеют более сложную структуру, чем их предшественники. Исчезает типичный нейропиль. Нейроны в ганглии занимают не только периферическое, но и центральное положение.
o концентрации ганглиев вокруг жизненно важных центров животного: головной конец, гонада, мышцы ноги (у двустворчатых и брюхоногих моллюсков).
 появление специализированных отростков нейронов. Развитие сложных воспринимающих чувствительных структур и моторных, двигательных элементов животного, потребовало более точной и адресной иннервации. С возникновением дендритов и аксонов дальнейшее функционирование нервной системы стало осуществляться по принципу рефлекса.
появление специализированных отростков нейронов. Развитие сложных воспринимающих чувствительных структур и моторных, двигательных элементов животного, потребовало более точной и адресной иннервации. С возникновением дендритов и аксонов дальнейшее функционирование нервной системы стало осуществляться по принципу рефлекса.
 цефализация. У высокоргнанизованных в эволюционном отношении беспозвоночных животных (насекомые, головоногие моллюски) ганглии сливаются с образованием общей массы наподобие головного мозга позвоночных животных (рис. 19).
цефализация. У высокоргнанизованных в эволюционном отношении беспозвоночных животных (насекомые, головоногие моллюски) ганглии сливаются с образованием общей массы наподобие головного мозга позвоночных животных (рис. 19).
5.3. Трубчатый тип нервной системы
У позвоночных животных в основе формирования нервной системы лежит нервная трубка, расположенная с дорсальной стороны эмбриона. Передний конец трубки обычно расширен и образует головной мозг. Задняя цилиндрическая часть есть ни что иное, как спинной мозг. Существует гипотеза, согласно которой у предков хордовых животных имелась продольная дорсальная полоса первичного чувствующего эпителия. Затем в ходе эволюционного развития она стала погружаться в эктодерму, сформировав сначала открытый желоб, а потом образовав замкнутую нервную трубку. Эту гипотезу подтверждают картины раннего эмбриогенеза позвоночных животных (рис.20).
 |
В ходе эволюционного развития позвоночных трубчатая нервная система претерпевает ряд изменений.
Во-первых, все дальнейшее развитие идет по пути цефализации – преимущественного развития головного мозга. Если у примитивно устроенного хордового животного – ланцетника – головной конец практически не развит, то уже у круглоротых имеется заметное утолщение нервной трубки в головном конце. Этот довольно примитивный головной мозг уже состоит из трех отделов: переднего, среднего и заднего. Передний отдел связан с развитием обоняния, средний – зрения, а задний с механорецепцией. У рыб выделяется также промежуточный мозг, достаточное развитие получает мозжечок. У амфибий значительно увеличивается передний мозг за счет развития полушарий, хорошо развит средний мозг, который у данной группы животных является высшим зрительным центром.
Во-вторых, у высокоорганизованных позвоночных возникает новый отдел головного мозга – кора больших полушарий (плащ конечного мозга). Эта структура все больше подчиняет себе рефлексы низших отделов мозга, осуществляет за ними контроль. Данный этап получает название кортикализации (от лат. cortex – кора). У рептилий появляется кора головного мозга. Развитие головного мозга млекопитающих характеризуется усилением развития новой коры, появляется Варолиев мост, совершенствуются структуры среднего и продолговатого мозга (рис. 21). У высших млекопитающих ассоциативные зоны коры являются высшим центром интегративной деятельности в ЦНС.
Таким образом, увеличение объема и усложнение структуры отделов головного мозга позвоночных тесно связаны с развитием сенсорных систем и интегративной деятельности. Постепенно, в зависимости от притока сенсорной информации, в существующих отделах мозга появляются филогенетически новые образования, которые берут под свой контроль все большее количества функций.
Следует, однако, отметить, что и у позвоночных, в том числе и у высших, сохраняются черты предшествующих эволюционных типов нервных систем: ганглионарной и диффузной.
Так в периферической нервной системе (соматической и вегетативной) чувствительные нейроны образуют ганглии (спинномозговые, симпатические и парасимпатические). При помощи ганглионарых структур в человеческом организме обеспечивается работа эволюционно древних (по сравнению с психикой) структур. Это, прежде всего, восприятие (чувствительность, рецепция) и автономная деятельность внутренних органов.
Также в нашем организме сохранились и признаки диффузной нервной системы. Она образует третий отдел вегетативной нервной системы – метасимпатическую нервную систему (напомним, первые два отдела: симпатическая и парасимпатическая). Данный отдел обеспечивает автономную работу полых внутренних органов. Нейроны метасимпатической нервной системы образуют микроганглионарные скопления внутри стенок органов, координируя их деятельность (например, перистальтические движения кишечника, обеспечивающие продвижение пищи). Процессы, протекающие в диффузной нервной системе, могут видоизменяться под влиянием симпатической и парасимпатической нервных систем.
Дата: 2018-11-18, просмотров: 467.