И. А. Кирсанова
Анатомия ЦНС
курс лекций
для студентов гуманитарных специальностей

Владивосток
2007
Министерство образования и науки Российской Федерации
Федеральное агентство по образованию
Дальневосточный государственный университет
Кафедра клеточной биологии
И. А. Кирсанова
Анатомия ЦНС
курс лекций
для студентов гуманитарных специальностей
Владивосток
Издательство Дальневосточного университета
2007
УДК 612.82(075.8)
ББК 28.706.3
К 43
Ответственный редактор
А.П. Анисимов, докт. биол. наук, профессор
Кирсанова И.А.
Анатомия ЦНС. Курс лекций.-
К 43 Владивосток: Изд–во Дальневост. ун–та, 2007. - с.
В учебном пособии рассматриваются строение и общие принципы организации нейрона и нервной ткани, показаны общие закономерности формирования нервной системы в эволюционном и индивидуальном развитии. Описывается строение и связи отделов центральной нервной системы человека. Особое внимание уделяется структурам мозга, обуславливающим психические функции.
Учебное пособие предназначено для студентов гуманитарных специальностей.
К 2007010000
180 (03) - 2007 ББК 28.706.3
© Кирсанова И.А.
© Издательство Дальневосточного университета, 2007
Предисловие
Анатомия центральной нервной системы (ЦНС) является базовой биологической дисциплиной при подготовке психологов и социальных работников. Фундаментальные данные о строении отделов ЦНС не изменялись в течение многих десятков и даже сотен лет. Вместе с тем активное развитие нейробиологии и нейрофизиологии в последние годы несколько изменило представление об устройстве и функции как нервных клеток, так и отдельных областей нервной системы. В ходе курса студенты получат знания об основных принципах организации нервной системы, познакомятся с закономерностями ее развития в онтогенезе с учетом возрастных особенностей, и в филогенезе – ее историческом, эволюционном развитии.
Для подготовки будущих психологов данная дисциплина является теоретической основой позволяющей понять законы поведения животных, законы становления и функционирования человеческой психики. Последующие дисциплины, усвоение которых опирается на данные «Анатомии ЦНС» - это физиология ЦНС, физиология высшей нервной деятельности и сенсорных систем, психофизиология, нейропсихология, психогенетика и др.
Главной задачей данного курса является формирование у студентов представления о многоуровневой организации нервной системы. К другим, не менее важным задачам относятся:
- создать у студентов представление о ЦНС, как об одной из интегративных систем организма,
- сформировать у студентов представление о нервной системе как материальном субстрате психической деятельности,
- научить студентов распознавать основные структуры мозга, объяснять их значение,
- применять знания об организации и основных принципах работы мозга в смежных дисциплинах.
Подробное описание морфологии ЦНС на субклеточном, клеточном, тканевом и системном уровнях с учетом приведенных иллюстраций облегчит усвоение теоретического материала. Определенную трудность для студентов представляет собой необходимость усвоения терминологии данной дисциплины. В настоящем пособии предпринята попытка представить материал достаточно подробно, с учетом академичности изучаемой дисциплины, в сочетании с его доступностью для студентов-гуманитариев. С той же целью в пособии приведен список научных терминов, которые помогут студентам при самостоятельной работе с использованием фундаментальных учебников по анатомии.
Лекционный курс рассчитан на 37 ч и состоит из шести разделов, включающих 15 тем. В первом разделе « Нервная ткань» рассматривается микроанатомическое строение ЦНС: нейроны и глия, а также строение нервов. Второй раздел «Развитие нервной ткани» содержит информацию о развитии нервной системы в филогенезе и онтогенезе. Третий раздел «Спинной мозг» и четвертый раздел «Головной мозг», посвящены изучению макроанатомии основных отделов ЦНС. Пятый раздел «Вегетативная нервная система» содержит данные об особенностях строения автономной нервной системы. В шестом разделе приведены общие сведения об эндокринной системе, затрагиваются вопросы взаимодействия двух регуляторных систем организма – нервной и гуморальной.
Введение
1.1. Содержание предмета «Анатомия ЦНС»
1.2. Методы
1.3. Задачи
1.4. Строение нервной системы
1.5. Анатомическая номенклатура
1.1. Содержание предмета «Анатомия ЦНС»
Классическая анатомия центральной нервной системы представляет собой раздел фундаментальной академической дисциплины - анатомии человека. Анатомия человека рассматривает форму и строение человеческого тела, всех его частей и органов в связи с их функцией, развитием и влиянием окружающей организм среды. Анатомия центральной нервной системы дает представление об устройстве и топографо-анатомических отношениях мозговых структур с учетом их микроскопического строения, с привлечением современных сведений о функциональном назначении каждой структуры.
Особенностью данного предмета является невозможность формирования правильных представлений об устройстве нервной системы без учета с одной стороны определенных взаимоотношений между организмом и окружающей его средой, с другой стороны - определенных взаимоотношений внутри самого организма. Сам организм человека представляет собой единое целое, сложный комплекс частей и органов, не просто механически соединенных, а находящихся в функциональной взаимосвязи, где всякое изменение в его любой части неизбежно отражается на деятельности всех остальных частей. Поэтому при изучении строения нервной системы необходимо учитывать ее двусторонние связи с другими анатомическими структурами. Важно и влияние химических факторов, которое лежит в основе взаимосвязи двух интегрирующих систем организма – нервной и гуморальной. Эти взаимоотношения достаточно сложны, даже если бы мы рассматривали организм вне связи с окружающей средой. Для человека отношения с окружающей средой еще более усложняются разнообразными социальными взаимоотношениями.
Необходимо помнить, что анатомические особенности нервной системы также определяются и особенностями развития - как индивидуального (онтогенез), регулируемого в основном эпигенетическими факторами – внешними и внутренними, так и эволюционного (филогенез), представляющего собой результат генетических преобразований.
В результате всего вышеперечисленного в рамках курса «Анатомия ЦНС» нам предстоит ответить на следующие вопросы:
 как устроена нервная система на микро- и макроскопическом уровнях
как устроена нервная система на микро- и макроскопическом уровнях
каким образом морфологические особенности нервной ткани определяют функции нервных структур
почему именно так организована нервная система человека
Методы
Название «анатомия» происходит от греческого слова anatemno - рассекаю. Это связано с тем, что первым методом анатомического исследования являлось рассечение трупа на части, или препарирование. Хороший анатомический препарат позволяет правильно определить и грамотно описать топографо-анатомические взаимоотношения отделов ЦНС на макроскопическом уровне.
Развитие клеточной биологии, техники микроскопического препарирования: световой, электронной, конфокальной микроскопии - дополнило данные макроанатомии сведениями о клеточном строении нервной системы, об ультраструктурных особенностях нейронов, глиальных клеток; о строении специализированных контактов нейронов – синапсов. Наука, родившаяся на стыке нейроанатомии, нейрофизиологии и нейрохомии, и расширившая наши познания в области организации субклеточных структур нервной ткани, их биофизических свойств, получила название нейробиологии.
Следует сказать несколько слов о методах, не являющихся непосредственно методами анатомии ЦНС, но, безусловно обогативших наши представления о ней. Современное развитие науки и техники позволяет изучать не только иссеченные (посмертные) препараты нервной ткани, появились методы дающие возможность прижизненного изучения мозговых структур. Сюда следует отнести основные методы медицинской диагностики: энцефалографию, эхоэнцефалоскопию, компьютерную томографию и др., дающие представление об изменении структуры и функций нервной ткани при изменении физической нагрузки и развитии заболеваний.
Задачи
Все вышеперечисленные методы позволяют решить следующие задачи:
изучить детали внешнего и внутреннего строения мозговых структур
установить уровни организации и интеграции отдельных нервных элементов и отделов ЦНС в организме
выявить роль различных структур нервной системы в жизнедеятельности животных и человека
Иными словами, все задачи и методы «Анатомии ЦНС» позволяют нам ответить на вопросы «как устроена нервная система организма, почему она устроена именно таким образом?». Ответ на вопрос, «каким образом это работает?» студенты получат, изучив следующий курс – «Физиологию ЦНС».
1.4. Строение нервной системы
Нервная система в соответствии с общей топографией организма разделена на два отдела: центральный и периферический. Центральный отдел составляют те части нервной системы, которые находятся внутри костных образований. Головной мозг – часть ЦНС, заключенная внутри черепа, спинной мозг – часть ЦНС, заключенная внутри позвоночника. К периферическому отделу относят те части нервной системы, которые лежат вне черепа или позвоночника – нервы и нервные узлы.
В соответствии с другой классификацией, в основе которой лежат функциональные особенности нервной регуляции, всю нервную систему (и центральную и периферическую) делят на два других отдела: соматический и вегетативный. Основные отличия этих отделов представим виде схемы:

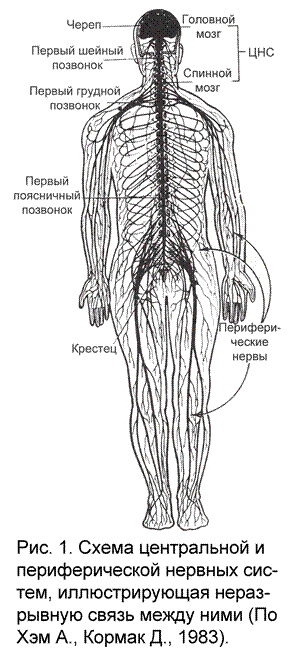 Нужно отчетливо представлять, что разделение нервной системы на соматическую и вегетативную, равно как и на центральную и периферическую носит условный характер, так как все отделы нервной системы анатомически и функционально связаны друг с другом, являются одним целым (рис. 1). Более того, между центрами головного и спинного мозга и регулируемыми ими органами существует не только прямая, но и обратная связь. Иными словами, из органов, деятельность которых изменяется под влиянием нервных импульсов, посылаемых мозговыми центрами, обратно в мозг поступает информация о характере этих изменений.
Нужно отчетливо представлять, что разделение нервной системы на соматическую и вегетативную, равно как и на центральную и периферическую носит условный характер, так как все отделы нервной системы анатомически и функционально связаны друг с другом, являются одним целым (рис. 1). Более того, между центрами головного и спинного мозга и регулируемыми ими органами существует не только прямая, но и обратная связь. Иными словами, из органов, деятельность которых изменяется под влиянием нервных импульсов, посылаемых мозговыми центрами, обратно в мозг поступает информация о характере этих изменений.
Нервная система образована специфической возбудимой тканью – нервной тканью. Основными клетками нервной ткани являются нервные и глиальные клетки.
Нервная клетка (нейрон) специализирована таким образом, что она способна приходить в состояние возбуждения:
принимать определенные формы сигналов
отвечать специальными сигналами
создавать специфические контакты с другими клетками.
Из-за этих свойств нейрон считается структурно-функциональной единицей нервной ткани.
Глиальные (греч. glia - клей) клетки выполняют многие функции в нервной системе, но никогда не проводят нервных импульсов. Различают несколько видов глиальных клеток, для них характерны опорная, трофическая, секреторная, защитная и некоторые другие функции.
Центральная нервная система - головной и спинной мозг – состоят из огромного количества нервных клеток, их отростков, а также из нейроглии, образующих серое и белое мозговое вещество. Серое вещество представляет собой скопление нервных клеток с отходящими от них отростками. Белое вещество – скопление нервных волокон. Нейроглия входит в состав как серого, так и белого вещества. В головном мозге, в разных его отделах серое вещество расположено неодинаково. В больших полушариях и в мозжечке серое вещество образует снаружи сплошной слой, называемый корой (cortex). Под корой находится белое вещество, а в нем – отдельные скопления серого вещества – ядра (nucleus). В других отделах головного мозга белое вещество расположено снаружи, а серое вещество в виде ядер внутри. Некоторые отделы состоят преимущественно из ядер. В спинном мозге белое вещество всегда снаружи, а серое внутри, также имеет ядерное строение. Ядра серого вещества играют роль различных центров головного и спинного мозга, регулируют функции органов (центр слюноотделения, центр дыхания, центр глотания и т.д.). Пучки нервных волокон белого вещества связывают одни отделы мозга с другими и выполняют проводниковую функцию.
Периферическая нервная система представлена черепными и спинномозговыми нервами и их окончаниями. Нервы состоят из пучков нервных волокон, окруженных общими оболочками. Многие нервы и их разветвления имеют помимо нервных волокон и нервные узлы – ганглии (ganglion). Они состоят из нейронов, отростки которых входят в состав данных нервов. Нервы дают ветви ко всем органам нашего тела, или как принято говорить иннервируют их. Нервные окончания могут быть образованы с чувствительными клетками, они получают название рецепторных (или афферентных), и с двигательными – эффекторные (или эфферентные). Сочетание рецепторных, нервных и эффекторных компонентов формируют разные по уровню сложности организации рефлекторные дуги.
Анатомическая номенклатура
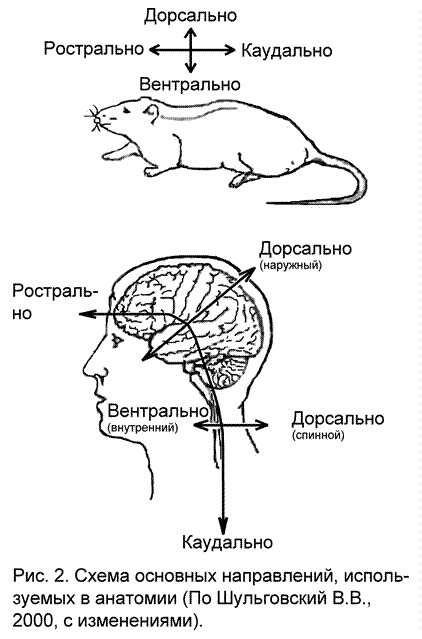
Для обозначения взаимного расположения структур внутри организма анатомы пользуются специальными терминами, основанными на латинских названиях (рис. 2; приложение 1):
плоскость, проходящая вдоль по середине тела, делящая его на правую и левую половины, называется сагиттальной (sagittalis)
плоскость, идущая также идущая вдоль по середине тела, но перпендикулярно сагиттальной, называется фронтальной (она параллельна лбу – frontis)
структуры, расположенные на спинной стороне тела называются дорсальными (dorsalis – спинной)
структуры, расположенные на брюшной стороне – вентральными (ventralis – брюшной)
структуры, лежащие по центру тела, близко к сагиттальной плоскости – медиальными (medialis – срединный)
структуры, лежащие сбоку от сагиттальной плоскости – латеральными (lateralis – боковой)
самые верхние точки структур называются апикальными (apicalis – верхушечный)
структуры, лежащие в основании – базальными (basalis)
направление к верхней части тела, к головной его части – ростральное (rostrum – клюв)
направление к нижней части тела – каудальное (caudalis – хвостовой)
По отношению к протяженным органам – конечностям, нервам и т.п. используются следующие термины:
проксимальный – близкий от начала отхождения (от туловища, от спинного или головного мозга)
дистальный - отдаленный
Раздел 1
НЕРВНАЯ ТКАНЬ
Клеточная мембрана
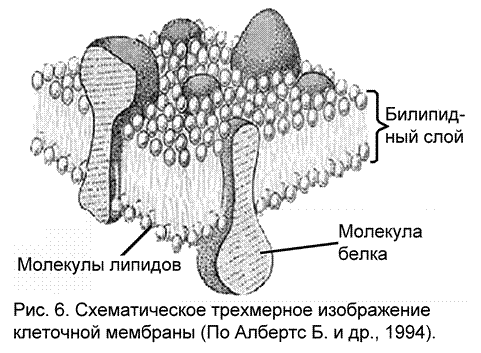
Основные свойства нервной клетки – раздражимость, возбудимость и проводимость определяются, как и у других электровозбудимых клеток (секреторных, мышечных), строением клеточной мембраны. В нервной ткани клеточная мембрана (рис. 6) может быть охарактеризована следующим образом:
с морфологической точки зрения – она состоит из нескольких слоев: два слоя жиров - фосфолипидов, пронизанных белковыми молекулами, снаружи мембраны расположен рецепторный белково-полисахаридный комплекс.
с физико-химической точки зрения – это сложная жидкая система макромолекул, в которой белки могут относительно свободно перемещаться: белковые молекулы могут менять степень своей погруженности в билипидный (bi – двойной, lipid – жир) слой клеточной мембраны, но всегда сохраняют контакт с окружающей средой с помощью полярной группы.
с функциональной точки зрения – это образование, которое окружает клетку, как самостоятельную живую единицу и является посредником ее контактов с окружающей средой, регулятором протекающих при этом обменных процессов. Белки клеточной мембраны выполняют следующие функции:
1) структурную
2) рецепторную - у белков наружной поверхности есть активный центр, который обладает сродством к различным веществам – гормонам, биологически активным веществам и др.
3)
 |
ферментативную - активируется под влиянием различных факторов
4) транспортную - полностью погруженные в билипидный слой белки образуют каналы, через которые проходят различные вещества. Обнаружены каналы для всех ионов, принимающих участие в возникновении процессов раздражения и возбуждения нейронов: K+, Na+, Ca2+, Cl-
Такое строение мембраны позволяет клетке, нейрону, избирательно проводить и накапливать внутри себя необходимые для процессов жизнедеятельности химические вещества (электролиты и неэлектролиты), а также избавляться от ненужных веществ.
Перенос веществ через мембрану осуществляется несколькими основными путями:
через билипидный слой. Барьерная роль клеточной мембраны заключается в ограничении свободной диффузии веществ. Модельные опыты на искусственных липидных мембранах показали, что они проницаемы для воды, газов, малых неполярных молекул жирорастворимых веществ, но совершенно не проницаемы для заряженных молекул (ионы) и для крупных незаряженных (сахара). Естественные мембраны так же ограничивают скорость проникновения низкомолекулярных соединений в клетку.
пассивным транспортом, или диффузией. При таком способе ионы перемещаются по белковым каналам в соответствии с градиентом концентрации, т.е. в направлении от более высокой концентрации веществ (ионов), к более низкой. Этот вид транспорта не требует никаких затрат энергии и продолжается до тех пор, пока не установится электрохимическое равновесие по обе стороны мембраны, а следовательно, пока не исчезнет градиент.
активным транспортом - осуществляется за счет ферментативной функции белков. Он всегда требует затрат энергии и направлен против градиента концентрации переносимых веществ. При помощи активного транспорта клетка получает из окружающей среды вещества, необходимые ей для процессов жизнедеятельности.
2.2.2. Цитоплазма и клеточные органеллы
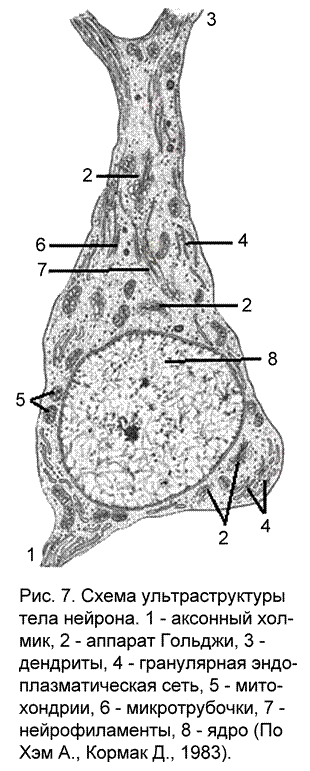
Любая клетка, в том числе и нервная, является единой системой, состоящей из множества закономерно связанных друг с другом элементов, представляющих собой определенное целостное образование, состоящее из сопряженных функциональных единиц – органелл или органоидов (рис. 7). Все, что находится внутри плазматической мембраны (кроме ядра и органелл) называется цитоплазмой. Это внутренняя среда клетки, отличающаяся высоким содержанием воды, белков, липидов, углеводов и солей. Она обеспечивает основные метаболические процессы в клетке, и ее перемещение называется цитоплазматическим потоком. Поскольку нейроны обладают довольно длинными отростками, объем которых может в десятки и сотни раз превышать объем тела нейрона, типы и формы перемещения в цитоплазме должны соответствовать функциональным требованиям. В зависимости от специализации отростков выделяют дендритный транспорт, аксонный (медленный, обеспечивающий рост, поддерживающий существование аксона и быстрый, который транспортирует компоненты, необходимые для работы синапсов) и ретроградный - направленный от периферии отростков к телу клетки.
Ядро нейрона, как и ядра других клеток, является местом хранения и реализации генетической информации. Вместилищем этой генетической информации являются молекулы ДНК, которые представляет собой чрезвычайно длинные линейные структуры, состоящую из двух взаимозакрученных полимерных цепей. Различные участки молекулы ДНК ответственны за синтез разных белков, одна молекула ДНК может определить синтез большого числа функционально и химически различных белков клетки. Однако синтез белков происходит отнюдь не в ядре, а в цитоплазме, не мелких частицах – рибосомах. Посредником в этом процессе являются молекулы РНК, синтезируемые в ядре, структура которых подобна структуре ДНК. Молекулы РНК переходят сквозь ядерную мембрану в цитоплазму и принимают непосредственное участие в синтезе белков.
 Вещество Ниссля. В световой микроскоп в нейронах, окрашенных по методу Ниссля (Франц Ниссль, немецкий невропатолог 19 в.) хорошо видны глыбки некоего клеточного вещества, расположенного в телах и крупных дендритах. Эти глыбки получили название телец Ниссля. В электронный микроскоп видно, что это вещество образовано гранулярной эндоплазматической сетью, содержащей многочисленные свободные и прикрепленные к мембранам сети рибосомы. Как уже было сказано – рибосомы являются местом синтеза белков, которые затем направляются в отростки (аксоны и дендриты) для замещения белков, использованных в метаболизме.
Вещество Ниссля. В световой микроскоп в нейронах, окрашенных по методу Ниссля (Франц Ниссль, немецкий невропатолог 19 в.) хорошо видны глыбки некоего клеточного вещества, расположенного в телах и крупных дендритах. Эти глыбки получили название телец Ниссля. В электронный микроскоп видно, что это вещество образовано гранулярной эндоплазматической сетью, содержащей многочисленные свободные и прикрепленные к мембранам сети рибосомы. Как уже было сказано – рибосомы являются местом синтеза белков, которые затем направляются в отростки (аксоны и дендриты) для замещения белков, использованных в метаболизме.
Участок тела нервной клетки, от которого отходит аксон, называемый аксонным холмиком (рис. 7, 1), относительно свободен от гранулярной эндоплазматической сети. Этот участок вместе с начальным безмиелиновым фрагментом аксона формируют инициальный сегмент (рис. 4а) – зону повышенной возбудимости в нервной клетке.
Другие клеточные органеллы в нейронах мало отличаются от органелл клеток ненервной природы. Имеется достаточно большое количество митохондрий, обеспечивающих клетку энергией, которая запасается в виде высокоэнергетических молекул (АТФ) и расходуемых клеткой по мере необходимости. Также развита сеть внутренних мембранных канальцев, свободных от рибосом, которые в зависимости от степени агрегации называются либо гладкой эндоплазматической сетью, либо аппаратом Гольджи (именно в телах нейронов в 1898 году Гольджи выявил аппарат, который назвал собственным именем). Здесь происходит синтез полисахаридов, липидов, идет дозревание белков или нейросекрета. Имеются в нейронах и типичные сетчатые клеточные образования – микротрубочки. Это тонкие опорные структуры белкового происхождения, помогающие нейрону сохранять свою форму и форму своих отростков. Здесь есть и толстые белковые нитчатые структуры - нейрофиламенты. Они принимают участие в транспорте веществ и органелл по отросткам нейронов.
Классификация нейронов
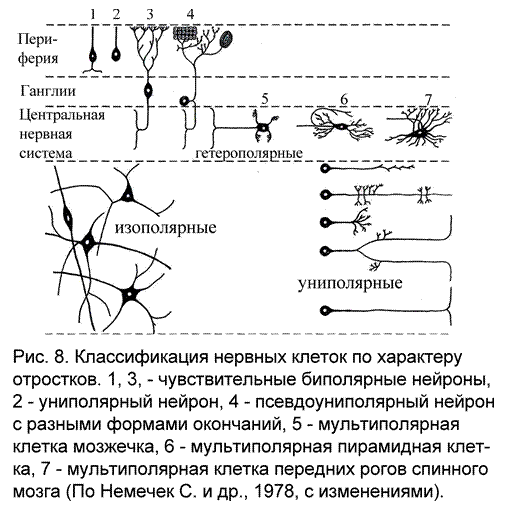 |
Наиболее часто встречающаяся классификация нейронов исходит из количества и характера отростков, отходящих от тела нервной клетки (рис. 8):
униполярные - имеющие один отросток, отходящий от тела клетки. Если этот отросток разделяется с образованием ветвей, специализирующихся как дендрит и аксон, то такой тип нейронов называют псевдоуниполярным.
биполярные – от тела клетки отходят два отростка: один аксон и один дендрит.
мультиполярные – нейроны, имеющие один аксон и много дендритов.
Если возможно отличить аксон от дендрита, то речь идет о гетерополярных нейронах. Обычным элементом нервной системы позвоночных является мультиполярный гетерополярный нейрон. В изополярных нейронах невозможно отличить дендрит от аксона. Такие нервные элементы характерны для низших беспозвоночных.
По функциональному признаку различают:
рецепторные (синонимы: сенсорные, афферентные, чувствительные, по положению в рефлекторной дуге – центростремительные) – это нервные клетки, по которым импульсы идут от рецепторов в ЦНС. Они делятся на первичные афферентные нейроны (их тела расположены в спинальных ганглиях, имеют непосредственную вязь с рецепторами) и вторичные афферентные нейроны (их тела лежат в стволе головного мозга, они не связаны с рецепторами, получают импульсы от других нейронов)
эфферентные (синонимы: двигательные, моторные, центробежные) – передающие импульсы из ЦНС к другим органам
вставочные (интернейроны, интегрирующие, модулирующие: фоновые, командные) - обеспечивают передачу импульсов с афферентных нейронов на эфферентные. Они составляют основную массу серого вещества головного мозга.
Нервные клетки из-за своей формы также могут получать специфические названия: пирамидные, грушевидные, веретеновидные, корзинчатые, звездчатые и др.
В нервной системе существуют особые нервные клетки – нейросекреторные. Они, как и все нейроны способны проводить и генерировать нервные импульсы, но их специфической особенностью является способность выработки и выделения наружу (т.е. секреции) биологически активных веществ. Нейросекреторные клетки является связующим элементом между двумя основными регуляторными системами организма – нервной и гуморальной. Аксоны нейросекреторных клеток характеризуются многочисленными расширениями, которые возникают в связи с временным накоплением нейросекрета. Самые крупные и гигантские расширения называются «телами Геринга». Из-за своей функциональной особенности, проявляющейся в экспорте нейросекрета, данные клетки содержат большое количество вещества Ниссля (белоксинтезирующий аппарат) и отличаются хорошо развитым аппаратом Гольджи (синтез липидов, полисахаридов, дозревание нейросекрета).
Синапсы
3.1. Общая характеристика синапсов
3.2. Электрический синапс
3.3. Химический синапс
3.4. Рефлекторная дуга
Электрический синапс
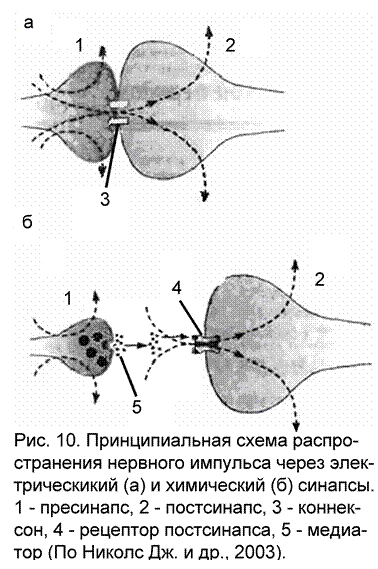 Существование электрических синапсов в природе подозревалось давно, но описаны они были только в 1957-59 гг. Такой тип клеточных контактов более характерен для беспозвоночных животных, у позвоночных он встречается реже. Отличительной чертой электрических синапсов является очень узкая синаптическая щель, а электрические взаимодействия возможны благодаря высокопроводящим (низкоомным) соединениям – щелевым контактам (рис. 10а). Щелевым контактом называется участок мембран двух клеток, в каждой из которых имеется скопление особых частиц, именуемых коннексонами. Они состоит из шести белковых субъединиц, образующих круг с внешним диаметром около 10 нм и внутренним диаметром 2 нм. На сопряженной клетке находится точно такая же структура. Совместно они пронизывают зазор между мембранами (2-3 нм). Внутри коннексонов имеется полость, через которую происходит перемещение ионов и мелких молекул между клетками. При распространении нервного импульса потери виде утечки тока через синаптическую щель в окружающую среду минимальны. Именно эта особенность позволила распознать в эксперименте электрические синапсы. Впервые это обнаружили Эд. Фершпен и Д. Поттер при исследовании синапсов речного рака. В стволе мозга млекопитающих также найдены области с электрическими синапсами: в среднем мозге, между телами клеток ядра тройничного нерва, в вестибулярном ядре Дейтерса между телами клеток и окончаниями аксонов, в продолговатом мозге между дендритами. Есть данные о присутствии электрических синапсов в колонках коры больших полушарий. Электрические синапсы позволяют синхронизировать активность групп нейронов, что дает возможность получать постоянные стереотипные реакции организма при многократных воздействиях.
Существование электрических синапсов в природе подозревалось давно, но описаны они были только в 1957-59 гг. Такой тип клеточных контактов более характерен для беспозвоночных животных, у позвоночных он встречается реже. Отличительной чертой электрических синапсов является очень узкая синаптическая щель, а электрические взаимодействия возможны благодаря высокопроводящим (низкоомным) соединениям – щелевым контактам (рис. 10а). Щелевым контактом называется участок мембран двух клеток, в каждой из которых имеется скопление особых частиц, именуемых коннексонами. Они состоит из шести белковых субъединиц, образующих круг с внешним диаметром около 10 нм и внутренним диаметром 2 нм. На сопряженной клетке находится точно такая же структура. Совместно они пронизывают зазор между мембранами (2-3 нм). Внутри коннексонов имеется полость, через которую происходит перемещение ионов и мелких молекул между клетками. При распространении нервного импульса потери виде утечки тока через синаптическую щель в окружающую среду минимальны. Именно эта особенность позволила распознать в эксперименте электрические синапсы. Впервые это обнаружили Эд. Фершпен и Д. Поттер при исследовании синапсов речного рака. В стволе мозга млекопитающих также найдены области с электрическими синапсами: в среднем мозге, между телами клеток ядра тройничного нерва, в вестибулярном ядре Дейтерса между телами клеток и окончаниями аксонов, в продолговатом мозге между дендритами. Есть данные о присутствии электрических синапсов в колонках коры больших полушарий. Электрические синапсы позволяют синхронизировать активность групп нейронов, что дает возможность получать постоянные стереотипные реакции организма при многократных воздействиях.
Химический синапс
В 1892 году Лэнгли предположил, что синаптическая передача в вегетативных ганглиях млекопитающих имеет химическую природу, а не электрическую. Примерно через 10 лет Эллиот обнаружил, что экстракт, полученный из надпочечников, — адреналин — производит на клетки такое же воздействие, как и стимуляция симпатических нервов. Это позволило предположить, что адреналин может секретироваться нейронами, и выделятся нервными окончаниями при проведении возбуждения. Однако лишь в 1921 году Леви поставил опыт, в котором была установлена химическая природа передачи в вегетативных синапсах между блуждающим нервом и сердцем. Он наполнял физиологическим раствором сосуды сердца лягушки и стимулировал блуждающий нерв, вызывая замедление сердцебиений. Когда жидкость из заторможенного стимуляцией сердца была перенесена на второе нестимулированное сердце, оно в результате такого воздействия также начинало биться медленнее. Было очевидно, что стимуляция блуждающего нерва вызывала освобождение в раствор тормозящего вещества. В последующих экспериментах Леви и его коллеги показали, что ацетилхолин полностью воспроизводил эффекты этого вещества.
Удивительно, что идея этого эксперимента пришла Леви во сне, он записал ее, но поутру не смог разобраться в том, что написал ночью. К счастью, сон повторился, и в этот раз Леви не стал рисковать: он помчался в лабораторию и поставил этот эксперимент. Вот как он вспоминает эту ночь: По серьезному размышлению, в холодных лучах утреннего света, я бы никогда не поставил этого эксперимента. То, что блуждающий нерв может освобождать тормозящее вещество, было очень маловероятно; еще менее вероятным было то, что химическое вещество, которое предположительно должно было действовать на коротком расстоянии между нервным окончанием и мышцей, освобождалось бы в таком большом количестве, что сохраняло бы эффективность после разведения в перфузирующей жидкости.
В начале 1930-х годов роль ацетилхолина в синаптической передаче в ганглиях вегетативной нервной системы была окончательно установлена Фельдбергом и его сотрудниками. Дейл, британский физиолог и фармаколог, установил роль ацетилхолина при передаче возбуждения в нервно--мышечном синапсе.
Принципиальным отличием химического синапса передача раздражения с пресинапса на постсинапс при помощи биологически активных веществ, называемых медиаторами (трансмиттерами). В связи с этим появляются и отличия в морфологии химического синапса (рис. 10б; приложение 2):
пресинаптическая область обязательно содержит везикулы (замкнутые в собственные мембраны полости), наполненные медиатором. Везикулы способны к перемещениям, когда они подходят к мембране нервного окончания, то их мембраны сливаются, а медиатор высыпается в синаптическую щель.
синаптическая щель имеется между пре- и постсинаптическими мембранами. Величина щели неодинакова в различных синапсах. Так, в приведенном ранее примере нервно-мышечного окончания, где медиатором является ацетилхолин, ширина синаптической щели достигает 30 нм. Пространство синаптической щели заполнено межклеточной жидкостью, а в нервно-мышечном окончании коллоидом (поэтому часто синаптическую щель в нервно-мышечном окончании рассматривают как базальную мембрану).
постсинаптическая мембрана содержит на своей поверхности рецепторы – белки, обладающие большим сродством к медиатору. Определенному типу медиатора соответствует определенный тип рецептора, например, в нервно-мышечном окончании – холинорецепторы, они распознают медиатор в окружающей среде и вступают с ним во взаимодействие. В результате этого взаимодействия происходят структурные изменения на постсинаптической мембране, в результате которых изменяется ионная проводимость через постсинапс. Это, в свою очередь, является причиной дальнейшего распространения нервного импульса (в случае, если медиатор оказывает тормозящий эффект, то развитию процессов торможения на постсинаптической мембране).
Кроме ацетилхолина в качестве медиатора могут выступать также:
аминокислоты: глутамат, глицин, аспартат
биогенные амины: дофамин, g-аминомасляная кислота (ГАМК), норадреналин, адреналин, серотонин, гистамин
производные пурина: АТФ, АДФ, АМФ, аденозин
пептиды: энкефалины, эндорфины, вещество Р и многие другие.
Синапсы и нейроны, в соответствии с видом выделяемого медиатора называются по типу медиатора с добавлением окончания –эргический: холинэргический, адренэргический, ГАМКэргический, гистаминэргический и т.п.
Химические синапсы наиболее распространены в ЦНС позвоночных. В настоящее время известно, что нейрон может синтезировать и выделять несколько медиаторов (сосуществующие медиаторы). Нейроны также обладают нейромедиаторной пластичностью – способностью менять основной медиатор в процессе развития.
Рефлекторная дуга
При помощи синаптических контактов нейроны соединяются в нейронные цепи, которые в свою очередь являются структурной основой рефлекторной деятельности организма. По своему положению в цепи и в соответствии с выполняемой работой, нейроны подразделяются на чувствительные (афферентные), вставочные (промежуточные, центральные) и эффекторные (эфферентные) (рис. 11).
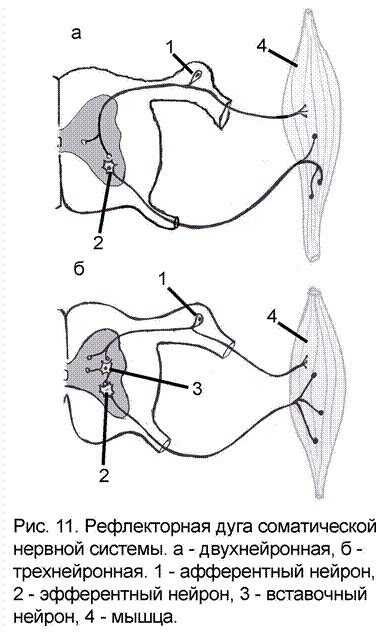 Перечисленные нейроны образуют рефлекторную дугу - путь, по которому проходят нервные импульсы от рецептора к исполнительному органу при осуществлении рефлекса. В ответ на рефлекторное воздействие от рабочего органа в ЦНС также поступают нервные импульсы, которые несут информацию о происходящих изменениях на периферии. Это звено работает по принципу обратной связи, осуществляя саморегуляцию деятельности организма.
Перечисленные нейроны образуют рефлекторную дугу - путь, по которому проходят нервные импульсы от рецептора к исполнительному органу при осуществлении рефлекса. В ответ на рефлекторное воздействие от рабочего органа в ЦНС также поступают нервные импульсы, которые несут информацию о происходящих изменениях на периферии. Это звено работает по принципу обратной связи, осуществляя саморегуляцию деятельности организма.
Принимая во внимание сигналы, поступающие от рабочего органа в ЦНС в ответ на рефлекторное воздействие, можно сказать, что рефлекторная дуга представляет собой сложный структурно-функциональный комплекс, замкнутый в кольцо. Компонентами такого рефлекторного комплекса являются:
рецептор – специализированная клетка, воспринимающая раздражение. Эта клетка переводит специфическую энергию раздражителя (механическую, термическую, химическую) в энергию нервного импульса.
афферентный путь – от рецептора к ЦНС
рефлекторный центр – совокупность нейронов, лежащих в ЦНС, в которых происходит обработка информации и формируется ответная реакция (образован вставочными нейронами)
эфферентный путь – от ЦНС на периферию
рабочий орган – мышца, железа
обратная связь – из органов, деятельность которых изменяется под влиянием эфферентных импульсов, посылаемых мозговыми центрами, обратно в мозг поступает информация о характере этих изменений (т.н. обратная афферентация). Различают два вида обратной связи: положительную – вызывает усиление ответной реакции, и отрицательную – вызывает торможение ответной реакции.
В зависимости от сложности строения рефлекторные дуги делят на простые и сложные.
Простые состоят из двух нейронов – чувствительного и двигательного, например, рефлекторная дуга коленного рефлекса. Теоретически в такой дуге может быть только один синапс, по другому она еще называется моносинаптической (приставка моно- означает «один»). На практике соответственно регистрируемому времени рефлекса, оказывается, что даже двухнейронная рефлекторная дуга имеет от 3 до 5 синапсов т.е. является полисинаптической (приставка поли- означает «много»). Простых рефлекторных дуг в организме человека немного, не более двух процентов от общего количества.
Сложные рефлекторные дуги содержат три и более нейронов – чувствительный, вставочный (от одного до десятков и сотен) и двигательный. Именно такие рефлекторные дуги наиболее распространены, а 90% нейронов ЦНС относятся к категории вставочных (приложение 3).
Также рефлекторные дуги, а, следовательно, и сами рефлексы можно классифицировать по следующим признакам:
в зависимости от того, где расположены рецептивные поля
o экстероцептивные – имеют рецепторы на внешней поверхности тела
o висцеро- или интероцептивные – имеют рецепторы во внутренних органах
o проприоцептивные – имеют рецепторы, регистрирующие движения (от мышц, связок, сухожилий)
по характеру ответной реакции
o соматические – проявляются в изменении функции с участием соматической нервной системы – произвольные движения.
o вегетативные – вызывают изменения в работе висцеральных органов.
в зависимости от того, где располагается рефлекторный центр данной дуги:
o спинальные – замыкаются на уровне спинного мозга
o бульбарные – на уровне продолговатого мозга
o мезэнцефальные – на уровне среднего мозга
o диэнцефальные – на уровне промежуточного мозга
o корковые – коры больших полушарий.
Глия. Строение нервов
4.1. Общая характеристика клеток глии
4.2. Типы глиальных клеток
4.2.1. Эпендима
4.2.2. Астроцитарная глия
4.2.3. Олигодендроглия
4.2.4. Микроглия
4.3. Строение нервов
Типы глиальных клеток
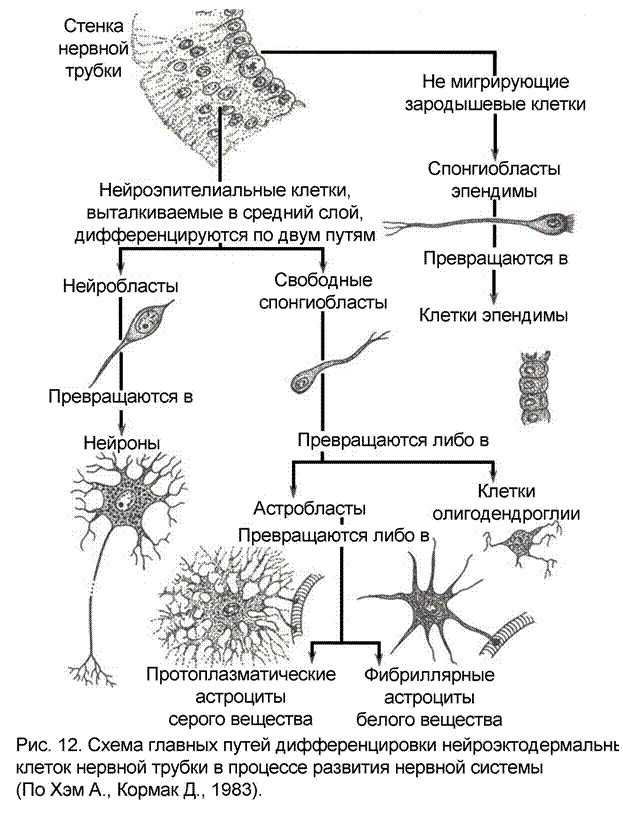
Когда Рудольф Вихров впервые ввел термин «глия» - он описал те элементы, которые позднее были названы астроглией. Название «глия» потом было перенесено и на другие клетки, позже открытые, которые не имели специализации нейронов, но находились в нервной ткани. Большая заслуга в вопросах классификации глии принадлежит испанской нейрогистологической школе, особенно Рамону-и-Кахалу и Рио-дель-Гортега. Морфологические различия отдельных типов клеток глии отвечают их разному назначению, разной специализации.
 |
4.2.1. Эпендима
С филогенетической и онтогенетической точки зрения эпендимная клетка является наиболее ранним элементом глии. У некоторых низших животных вся глиальная популяция нервной системы целиком состоит только из этих клеток. У высших позвоночных они образуют лишь выстилку полостей ЦНС, которая по своему строению напоминает цилиндрический эпителий и называется эпендимой.
Обращенная в полость поверхность эпендимной клетки покрыта короткими пальцеобразными отростками клеточной мембраны – микроворсинками и ресничками. Координированное движение ресничек (до 6 раз в секунду) является одним из механизмов, приводящих в движение жидкую среду полостей и желудочков ЦНС, которая по-другому называется ликвором. На боковых поверхностях эпендимных клеток находятся плотные щелевые контакты, которые укрепляют слой клеток, создавая сплошную структуру наподобие эпителиальной. С базальной стороны эпендимной клетки часто формируется отросток, контактирующий с другими клетками глии. Данный тип глиальных клеток выполняет три функции, осуществляемые в индивидуальном развитии более или менее последовательно.
Первая функция – пролиферативная – камбиальный резерв клеток зародышевой нервной трубки
Вторая функция – опорная. Когда стенка нервной трубки утолщается, выстилающие ее эпендимные клетки дают длинные отростки, которые достигают ее наружной поверхности и какое-то время способствуют образованию наружной мембраны, окружающей нервную трубку.
Третья функция – формирование непрерывной эпителиальной выстилки желудочков мозга. Эта выстилка сохраняется и в центральном канале спинного мозга. В некоторых участках желудочков эпендима участвует в формировании сосудистых сплетений мозга.
Астроцитарная глия
Распределение астроцитов в ЦНС равномерно, однако в делом веществе они встречаются реже, чем в сером. Ядра астроцитов крупнее ядер эпендимы, они светлее, часто имеют сложную форму (полиморфны). Для астроцитов характерно большое количество отростков, которые лучеобразно отходят от тела клетки. В астроцитах очень много фибриллярных структур, отсюда и их основная функция – опорная.
Вообще все астроциты делят на две большие категории – фибриллярные и протоплазматические (рис. 12). Фибриллярные астроциты содержат большое количество пучков фибрилл, расположены эти клетки в основном в белом веществе. Долгое время придерживались мнения, что только фибриллярные астроциты содержат фибриллы, но с помощью электронного микроскопа такие пучки были выявлены и в протоплазматических астроцитах. Последние имеют ветвящиеся цитоплазматические отростки, отходящие от тела клетки наподобие зарослей кустарника. Их отростки короче и ветви более многочисленны, чем у фибриллярных астроцитов.
Астроциты пронизывают всю ЦНС, контактируют между собой и с рецепторной частью нейронных синапсов. Астроциты составляют четвертую часть популяции глиальных клеток. По своей конфигурации астроциты полностью зависят от системы нейронов, которые они окружают. Аксоны нейронов часто бывают окутаны цитоплазмой астроцита. Астроциты также располагаются между капиллярами кровеносного русла и телами нейронов, и осуществляют транспорт веществ из крови в нейроны и обратно. Кроме того, астроглия связывает с кровеносным руслом спинномозговую жидкость.
Поскольку иногда встречаются дегенерирующие астроциты, возможно, что дегенерация находится в равновесии с образованием новых астроцитов, что указывает на возможность медленного обновления популяции этих клеток. Таким образом, основные функции астроцитарной глии:
опорно-механическая
трофическая
изоляционная
Олигодендроглия
Олигодендроциты имеют тоже происхождение, что и астроциты. По размерам они меньше, чем астроциты и имеют меньше отростков. Олигодендроциты находятся как в сером веществе (где представляют собой большую часть перинейрональных элементов) и так и в белом веществе (где ответственны за образование миелина, эти олигодендроциты обладают длинными отростками). Одним из типов олигодендроглии являются Шванновские клетки. Эти клетки располагаются в периферической нервной системе, способны спирально обертывать своей цитоплазмой аксоны нейронов и формировать миелиновые оболочки. Те олигодендроциты, которые находятся в сером веществе ЦНС, располагаются настолько плотно прилегая к телам нейронов, что получили название клеток-сателлитов. Основные функции олигодендроцитов:
изоляционная – шванновские клетки полностью ограничивают все нейрональные части периферической нервной системы от соединительной ткани
синтетическая – шванновские клетки берут на себя задачу создания белка миелина и обеспечения его структурной и функциональной целостности. Миелинизация в эмбриональном онтогенезе наступает достаточно поздно и протекает неделями и месяцами и после рождения. Основная функция – миелин позволяет увеличить скорость проведения нервных импульсов.
обмен веществ прилегающего нейрона – общая функция всех олигодендроцитов (и сателлитов и шванновских)
Микроглия
Клетки микроглии происходят из другого зародышевого листка – из мезодермы и играют большую роль при патологических состояниях. В мозг они прорастают на поздних стадиях эмбриогенеза, скапливаясь в тех местах, где мягкая мозговая оболочка непосредственно прилегает к белому веществу. Отсюда они распространяются по всей ткани ЦНС.
Некоторые авторы, например Артур Хэм и Дэвид Кормак считают что «… было бы проще, если бы микроглию совсем не причисляли к нейроглии, поскольку она не развивается из эпителия нервной трубки (как другие клетки мозговой ткани) и не выполняет поддерживающей функции».
Микроглия (как видно из названия) является наименьшим глиальным элементом в ЦНС. В количественном отношении микроглиальных клеток меньше чем астроцитов и олигодендроцитов. У этих клеток нет преимущественной локализации в нервной ткани, но в сером веществе их больше, чем в белом веществе. Часто они встречаются в виде сателлитов вокруг нервных клеток и сосудов. Цитоплазма этих клеток окружает ядро лишь узкой полоской. От тела клетки отходят многочисленные отростки, которые богато ветвятся.
Микроглия отличается повышенной подвижностью и способностью принимать и перерабатывать метаболиты (продукты жизнедеятельности других клеток). Микроглиальные элементы активизируются при болезненных процессах, сопровождающихся распадом нервной ткани. Основная функция микроглии – способность, подобно другим тканевым макрофагам, фагоцитировать (переваривать, утилизировать) гибнущие клетки ЦНС. Однако, как показали исследования, и в нормальных условиях микроглия сохраняет свою функциональную активность. Происходит постоянный обмен между нервной тканью, микроглией и кровью.
Строение нервов
Отростки нейронов, покрытые оболочками, называют нервными волокнами. Как мы уже упоминали, в зависимости от степени окруженности олигодендроцитами они бывают безмякотными (безмиелиновыми) и мякотными (миелиновыми). Пучки нервных волокон, окруженные соединительнотканными оболочками, называют нервами или нервными стволами. В составе некоторых нервов встречаются одиночные нервные клетки и мелкие ганглии или узлы (скопления нейронов). Нервы подразделяют на:
черепные – 12 пар
спинномозговые – 31 пара
Все нервы и их разветвления (вместе с концевыми аппаратами – рецепторами и эффекторами) составляют периферическую нервную систему. Посредством нервов и их разветвлений осуществляется связь ЦНС с органами, объединяются системы органов и осуществляется целостность организма.
Нервы являются довольно прочными структурами в отличие от тканей головного или спинного мозга. Это объясняется тем, что соединительнотканная оболочка нервов представляете собой трубчатую структуру трех порядков (рис. 13):
эпиневрий – наружная оболочка нервного ствола. Представлена рыхлой неоформленной соединительной тканью, богата коллагеновыми волокнами. Содержит фибробласты, жировые клетки, а также кровеносные и лимфатические сосуды
периневрий – тонкие прослойки соединительной ткани, разделяющие нерв на отдельные нервные пучки. Также содержит кровеносные и лимфатические сосуды.
эндоневрий – разделяет нервный пучок на отдельные нервные волокна.
В нервах небольшого диаметра эпиневрий отсутствует. Небольшие нервы состоят из периневрия, содержащего эндоневральные трубки, окружающие отдельные нервные волокна.
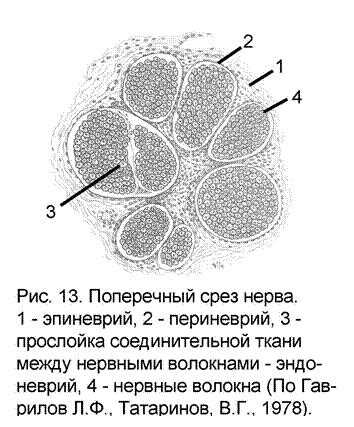 Такие же оболочки – эпи-, пери- и эндоневрий присутствуют и в цереброспинальных ганглиях.
Такие же оболочки – эпи-, пери- и эндоневрий присутствуют и в цереброспинальных ганглиях.
Нервы в зависимости от состава их волокон подразделяются на:
чувствительные – содержат центростремительные волокна
двигательные – центробежные волокна
смешанные – оба вида волокон
Большинство периферических нервов относится к смешанному типу. Ближе к дистальному концу нерва афферентные и эфферентные волокна рассортировываются, и отдельные нервные пучки становятся преимущественно чувствительными или двигательными. Число и диаметр нервных волокон в пучке варьируют. В дистальных частях некоторых нервов имеется больше волокон, чем в проксимальных частях. Увеличение числа волокон в нерве связано с их ветвлением.
Раздел 2
РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ
Онтогенез нервной ткани

Период внутриутробного развития начинается с момента слияния мужских и женских половых клеток и образования зиготы. Зигота последовательно делится, образуя шаровидную бластулу. На стадии бластулы идет дальнейшее дробление и образование первичной полости – бластоцеля.
Затем начинается процесс гаструляции, в ходе которого происходит перемещение клеток с образованием двухслойного зародыша. Наружный слой клеток называется эктодерма, внутренний – энтодерма. Внутри образуется полость первичной кишки – гастроцель. На этом заканчивается стадия гаструлы.
Зачаток нервной системы начинает развиваться уже в конце стадии гаструлы. На следующей стадии - нейрулы - образуются нервная трубка, хорда и сомиты (рис. 22).
 Процесс, благодаря которому осуществляется формирование нервной пластинки и превращение ее в нервную трубку, называется нейруляцией. Нейроны и глиальные клетки в ЦНС образуются в результате деления клеток-предшественников внутренней зоны нервной трубки. Нейроны мигрируют от внутренней поверхности нервной трубки и образуют серое вещество нервной системы взрослых позвоночных. Некоторые из клеток, расположенные по краям нервных валиков, остаются между нервной трубкой и вышележащей эктодермой. Эти клетки формируют нервный гребень. Клетки нервного гребня образуют чувствительную часть периферической нервной системы (рис. 23).
Процесс, благодаря которому осуществляется формирование нервной пластинки и превращение ее в нервную трубку, называется нейруляцией. Нейроны и глиальные клетки в ЦНС образуются в результате деления клеток-предшественников внутренней зоны нервной трубки. Нейроны мигрируют от внутренней поверхности нервной трубки и образуют серое вещество нервной системы взрослых позвоночных. Некоторые из клеток, расположенные по краям нервных валиков, остаются между нервной трубкой и вышележащей эктодермой. Эти клетки формируют нервный гребень. Клетки нервного гребня образуют чувствительную часть периферической нервной системы (рис. 23).
В пределах каждой из областей в развивающейся нервной системе судьба клеток в значительной степени зависит от их расположения и клеточного окружения. Фенотип, в который дифференцируются клетки нервного гребня, определяется сигналами, поступающими от соседних клеток. Таким образом, если клетки нервного гребня были пересажены в раннем возрасте, они будут дифференцироваться в соответствии со своим новым расположением.
 |
Уже на 3 - 4-й неделе выделяются две области нервной трубки: дорсальная (крыловидная пластинка) и вентральная (базальная пластинка). Из крыловидной пластинки развиваются чувствительные и ассоциативные элементы нервной системы, из базальной - моторные. Структуры переднего мозга у человека целиком развиваются из крыловидной пластинки. К концу 4-й недели головной конец нервной трубки расширяется, и из него начинает развиваться головной мозг, а из оставшейся части - спинной мозг. На этой стадии головной мозг представлен специализированными вздутиями – тремя первичными мозговыми пузырями (рис. 24). Из каждого пузыря развивается один из трех отделов мозга – передний, средний и задний. Во время сворачивания нервной трубки некоторые нервные клетки остаются вне ее, из них формируется глазной гребень. Вскоре после формирования трех первичных пузырей отмечаются первые признаки формирования глаз.
Затем наступает этап целой серии изгибов, которые предстоит претерпеть растущему мозгу, прежде чем он примет свои взрослые очертания. Изгибы разграничивают основные структурные единицы, и разделяют широкие внутренние полости, которые потом станут желудочками мозга.
В полтора месяца большой пузырь переднего мозга подразделяется на конечный мозг, из которого позже разовьется кора больших полушарий, и промежуточный мозг, из которого будут сформированы таламус и гипоталамус. Эти ранние стадии развития головного мозга напоминают соответствующие стадии формирования менее сложного мозга позвоночных. К двум месяцам уже образован основной (среднемозговой) изгиб головного мозга: передний мозг и промежуточный мозг загибаются вперед и вниз под прямым углом к продольной оси нервной трубки. Позже формируются еще два изгиба: шейный и мостовой. В этот же период первый и третий мозговые пузыри разделяются дополнительными бороздами на вторичные пузыри, при этом появляется 5 мозговых пузырей. Из первого пузыря образуются большие полушария головного мозга, из второго - промежуточный мозг. Из оставшихся пузырей формируются мозговой ствол и мозжечок. В течение 5 - 10-й недели развития начинается рост и дифференцировка конечного мозга: образуются кора и подкорковые структуры. На этой стадии развития появляются мозговые оболочки, формируются ганглии нервной периферической вегетативной системы. Спинной мозг приобретает окончательное строение.
 Конечный мозг в этот период проходит три стадии развития. На первой стадии развития появляются обонятельные доли мозга, гиппокамп, лимбическая система. Вторая стадия связана с ростом базальных ганглиев. Третья стадия развития конечного мозга включает формирование новой коры больших полушарий - неокортекса. Когда у приматов скорость роста неокортекса достигает максимума (ок. 250 тыс. клеток в минуту), поверхность его начинает образовывать извилины, которые позволяют увеличить объем корковой ткани без увеличения общих размеров мозга.
Конечный мозг в этот период проходит три стадии развития. На первой стадии развития появляются обонятельные доли мозга, гиппокамп, лимбическая система. Вторая стадия связана с ростом базальных ганглиев. Третья стадия развития конечного мозга включает формирование новой коры больших полушарий - неокортекса. Когда у приматов скорость роста неокортекса достигает максимума (ок. 250 тыс. клеток в минуту), поверхность его начинает образовывать извилины, которые позволяют увеличить объем корковой ткани без увеличения общих размеров мозга.
В следующие 10 - 20 недель беременности завершается формирование всех отделов головного мозга, идет процесс дифференцировки мозговых структур, который заканчивается только с наступлением половозрелости (рис. 25). Полушария становятся самой большой частью головного мозга. Выделяются основные доли (лобная, теменная, височная и затылочная), образуются извилины и борозды больших полушарий. В спинном мозге в шейном и поясничном отделах формируются утолщения, связанные с иннервацией соответствующих поясов конечностей. Окончательный вид приобретает мозжечок. В последние месяцы беременности начинается миелинизация (покрытие нервных волокон специальными чехлами) нервных волокон, которая заканчивается уже после рождения.
В процессе эмбрионального развития головного мозга полости мозговых пузырей видоизменяются и превращаются в систему мозговых желудочков, которые сохраняют связь с полостью спинномозгового канала. Центральные полости больших полушарий головного мозга образуют боковые желудочки довольно сложной формы. Боковые желудочки соединяются с полостью промежуточного мозга, которая является III желудочком. Через специальный проток (сильвиев водопровод) III желудочек соединяется с IV желудочком; IV желудочек образует полость заднего мозга и переходит в спинномозговой канал.
Раздел 3
СПИННОЙ МОЗГ
Оболочки мозга
Головной и спинной мозг защищены не только костным футляром (черепом и позвоночником), но и тремя соединительнотканными оболочками:
Мягкая мозговая оболочка (pia mater spinalis) – самая внутренняя, непосредственно прилегает к головному и спинному мозгу
Паутинная мозговая оболочка (tunica arachnoidea) – средняя оболочка
Твердая мозговая оболочка (dura mater spinalis) – наружная
Мягкая мозговая оболочка. Как можно судить по названию (pia – нежный, тонкий) – это оболочка тонкая. Она содержит тонкие переплетающиеся пучки соединительнотканных волокон, единичные фибробласты и макрофаги и много кровеносных сосудов, распределенных по всей поверхности мозга. Разветвления сосудов внедряются в спинной (головной) мозг. Она имеет два слоя: внутренний, сросшийся со спинным мозгом, наружный.
Паутинная оболочка. Средний слой мозговых оболочек называют паутинным, так как он отделен от мягкой оболочки (и в тоже время соединен с нею) паутинообразной сетью тонких соединительнотканных перегородок. Между мягкой и паутинной оболочками находится подпаутинное (субарахноидальное лимфатическое) пространство, заполненное цереброспинальной жидкостью. На поверхности головного мозга имеется много борозд и извилин. Мягкая оболочка повторяет ход борозд и извилин, вплотную покрывая их поверхность. А перепончатая часть паутинной оболочки напротив, не заходит туда (за исключением некоторых особенно крупных борозд). Потому над бороздами остается больше места для цереброспинальной жидкости, чем в других участках. Также имеются участки, называемые цистернами, где поверхность мозга и покрывающая ее паутинная оболочка разделены значительными расстоянием, так что здесь перемещается большое количество цереброспинальной жидкости.
Твердая мозговая оболочка. Эта наружная оболочка имеет жесткую консистенцию, так как состоит из плотной соединительной ткани. Количество соединительнотканных волокон здесь очень высоко. Имеются определенные различия в структуре твердой мозговой оболочки позвоночного канала и черепа. В позвоночном канале она лежит относительно свободно. Пространство между ее внутренней поверхностью и наружной поверхностью паутинной оболочки называется субдуральным (обычно содержит небольшое количество жидкости, которая не является цереброспинальной). Снаружи от твердой мозговой оболочки спинного мозга находится эпидуральное (экстрадуральное) пространство, заполненное рыхлой соединительной тканью, в которой имеется некоторое количество жировых клеток и многочисленные вены. Внутренняя надкостница позвоночника, выстилающая позвоночный канал служит наружной границей эпидурального пространства. В черепе нет эпидурального пространства, так как здесь твердая оболочка сливается с внутренней надкостницей черепных костей. Твердая оболочка содержит меньше сосудов, чем надкостница. Несмотря на срастание надкостницы, и твердой оболочки головного мозга, существуют участки, где они разделены. Обычно это участки крупных борозд, внутрь которых проникает твердая оболочка. Вдоль линии, от которой оболочка переходит в борозду, между складками твердой оболочки, может образовываться полость. Такие полости заполнены венозной кровью и называются венозными синусами и венозными лакунами.
Гематоэнцефалический барьер. Клинические и экспериментальные данные указывали на существование гематоэнцефалического барьера еще до того, как электронно-микроскопические исследования позволили установить его структурную основу. Оказалось, например, что некоторые лекарственные препараты при внутривенном введении не попадают из крови в головной мозг, хотя могут переходить в другие ткани. Кроме того, некоторые красители, обычно переходящие из крови в другие части тела, не обнаруживались в сером веществе, что, несомненно, свидетельствовало о существовании барьера между кровью и мозгом. На основании электронно-микроскопических исследований показано, что барьер образован:
непрерывным эндотелием капилляров (эндотелий – внутренний эпителиальный слой стенки сосудов), клетки которого соединены обширными плотными контактами, которые полностью исключают возможность существования больших щелей между клетками.
очень плотной базальной мембраной, окружающей капилляры.
Однако газы и другие мелкие молекулы, необходимые для питания нейронов и глиальных клеток, а также вирусы и некоторые споры грибов (вызывающие заболевания мозга) относительно легко диффундируют через этот барьер.
Диффузия – это облегченный вариант транспорта через полупроницаемые мембраны. Не требует затрат энергии, осуществляется по градиенту концентрации, т.е. в направлении от зоны с повышенным содержанием веществ, в зону с пониженным содержанием веществ.
Раздел 4
ГОЛОВНОЙ МОЗГ
Обзор головного мозга
9.1. Общая характеристика головного мозга
9.2. Основание мозга
9.3. Черепномозговые нервы
9.4. Сагиттальный срез головного мозга
Основание мозга
Нижняя поверхность (рис. 32), или основание мозга (basis cerebri) позволяют рассмотреть следующие анатомические образования.
Передний отдел основания мозга представлен лобными полушариями. Здесь имеется глубокая вертикальная щель (fissure longitudinalis cerebri , рис. 32,1), она продолжается до
перекреста зрительных нервов (ciasma opticus , рис.32,2). Спереди от перекреста находятся зрительные нервы (n . opticus , рис. 32,3 ), а сзади – зрительные тракты (tractus opticus , рис. 32,4).
По бокам от перекрестов и зрительных нервов лежит серое поле, усеянное мелкими (и более крупными) отверстиями – передняя продырявленная субстанция - (substantia perforate anterior , рис.32,5). Переднюю границу этого поля представляет обонятельный треугольник (trigonum olfactorium , рис.32,6), который ограничен наружной и внутренней обонятельными полосками (striae olfactoriae medialis et lateralis , рис. 32,7). От вершины обонятельного треугольника тянется узкая полоска – обонятельный тракт (tractus olfactory , рис. 32,8), заканчивающийся обонятельной луковицей (bulbus olfactorius , рис. 32,9).
 |
Позади перекреста зрительных нервов возвышается серый бугор (tuber cinereum , рис. 32,10), который вытягивается в отросток, напоминающий воронку (infundibulum, рис. 32,11). С его помощью основание мозга соединяется с гипофизом (hypophysis , рис.32,12). Слева и справа от серого бугра находятся ножки мозга (crus cerebri , рис.32,13), а позади него возвышаются два белых округлых образования – сосцевидные тела (corpora mamillaria , рис. 31,14).
Между ножками мозга кзади от сосцевидных тел лежит межножковая ямка (fossa interpeduncularis , рис. 32,15). Дно этой ямки усеяно многочисленными отверстиями - это задняя продырявленная субстанция (substantia perforate posterior). Она со стороны ножек ограничена бороздой. Здесь выходят волокна III-ей пары черепномозговых нервов – глазодвигательных (n . oculomotorius , рис. 32,16). Снаружи ножки мозга огибает IV пара нервов – нитевидный блоковый (n . trochlearis , рис. 32,17).
Позади этих образований находится поперечно расположенный валик – мост (pons Varoli , рис. 32,18). Посередине его видна уплощенная срединная борозда (sulcus basilaris , рис. 32,19). Кнаружи мост суживается, и затем, направляясь в стороны и назад, он погружается в мозжечок, формируя его средние ножки (pedunculus cerebellaris medius , рис. 32,20). У переднего края Варолиева моста на границе со средними ножками мозжечка локализуется V пара черепномозговых нервов – тройничных (n . trigeminus , рис. 32,21).
Кзади от моста располагается конусовидное образование – продолговатый мозг (medulla oblongata , рис. 32, 22). По средней линии продолговатого мозга проходит передняя срединная щель (fissura mediana anterior , рис. 32,23), ограниченная с каждой стороны белым валиком, или пирамидой (pyramis , рис. 32,24). Кнаружи от пирамиды лежит продолговатое возвышение, или олива (oliva , рис.32,25). На границе заднего края моста располагается место выхода VI пары черепномозговых нервов - отводящих (n . abducens , рис. 32,26). Снаружи от отводящего нерва и спереди от оливы на основании мозга имеются корешки VII пары – лицевого нерва (n . facialis , рис. 32,27) и VIII пары – преддверноулиткового (n . vestibulocochlearis , рис. 32, 28). Позади оливы определяются места выхода IX пары – языкоглоточного (n . glossopharyngeus , рис. 32,29), X пары – блуждающего (n . vagus , рис. 32,30), XI пары – добавочного (n . anccessorius , рис. 32,31) XII пара – подъязычный (n . hypoglossus , рис. 32,32) нерв появляется на основании мозга между пирамидой и оливой.
Продолговатый мозг перекрывает среднюю часть мозжечка и лежит в широкой его борозде, называемой долинкой мозжечка (vallecula cerebelli).
Вентральная поверхность мозжечка является резко выпуклой. Глубокая задняя срединная борозда (fissura medina posterior) разделяет полушария мозжечка (hemispheria cerebelli), которые покрыты многочисленными узкими извилинами, идущими более или менее параллельно.
Черепные нервы
Черепые нервы (nervi craniales) различаются по функциям, а, следовательно, и по составу нервных волокон и области иннервации.
I; II и VIII пары – чувствительные
V; VII; IX и X пары – смешанные
III; IV; VI; XI и XII пары – двигательные
Обонятельный и зрительный отличаются от других тем, что они являются производными головного мозга – образовались путем выпячивания из мозговых пузырей и в отличие от других чувствительных нервов не имеют ганглиев. Они состоят из аксонов нейронов, расположенных на периферии – в органе обоняния и органе зрения.
Смешанные по функциям черепые нервы по строению и составу волокна сходны со спинномозговыми нервами.
o Их чувствительная часть имеет узлы, аналогичные спинномозговым ганглиям. Дендриты нейронов этих узлов соединяются с рецепторами периферических органов, а аксоны следуют в ствол мозга к чувствительным ядрам, аналогичным ядрам передних рогов спинного мозга.
o Двигательная часть смешанных черепых нервов (и двигательные черепномозговые нервы) состоит из аксонов нервных клеток двигательных ядер ствола мозга.
В составе III– двигательных; VII; IX и X – смешанных пар нервов наряду с другими волокнами проходят парасимпатические волокна.
Обонятельные нервы (I пара) – по функции чувствительные, состоят из нервных волокон, являющихся центральными отростками обонятельных клеток, расположенных в органе обоняния. Эти волокна образуют 15-20 тонких обонятельных нитей, которые выходят из органа обоняния и через решетчатую кость проникают в полость черепа, где подходят к обонятельной луковице, образуя синапсы с ее клетками.
Обонятельные луковицы, обонятельные тракты, треугольники с передними продырявленным веществом, - это периферический отдел обонятельного мозга. По нему передается информация в центральный отдел, который представлен сводчатой извилиной, крючком, гиппокампом и пограничными с ним образованиями.
Зрительные нервы ( II пара) – по функции чувствительные. Состоят из нервных волокон т.н. ганглиозных клеток сетчатки глаза. Из глазницы через зрительный канал нерв проходит в полость черепа, где сразу образует частичный перекрест с нервом противоположной стороны и продолжается в зрительный тракт – все это периферический отдел зрительного анализатора.
В связи с тем, что на противоположную сторону переходит только медиальная половина нерва – правый зрительный тракт соединяет нервные волокна от правых половин глазных яблок, а левый – от левых.
Зрительные тракты подходят к подкорковым зрительным центрам – ядрам верхних холмиков крыши среднего мозга, латеральных коленчатых тел и подушек таламуса. Ядра верхних холмиков связаны с ядрами глазодвигательного нерва (через него осуществляется зрачковый рефлекс) и с ядрами передних рогов спинного мозга (осуществляется ориентировочный рефлекс на внезапные световые раздражения).
От ядер латеральных коленчатых тел и подушек таламуса нервные волокна в составе белого вещества полушарий следуют в кору затылочных долей (зрительная кора) – это центральный отдел зрительного анализатора.
Глазодвигательный нерв ( III пара) – по функции двигательный. Состоит из двигательных соматических волокон и эфферентных парасимпатических нервных волокон. Эти волокна составляют аксоны нейронов ядер нерва. Различают двигательные ядра и добавочное парасимпатическое ядро. Они находятся в ножке мозга на уровне верхних холмиков крыши среднего мозга. Нерв выходит из полости черепа через верхнюю глазничную щель в глазницу и делится на две части – верхнюю и нижнюю. Двигательные соматические волокна этих ветвей иннервируют поперечнополосатые мышцы: мышцу глазного яблока и мышцу, поднимающую верхнее веко. Парасимпатические волокна иннервируют гладкие мышцы: мышцу, суживающую зрачок и ресничную.
Блоковый нерв ( IV пара) – по функции двигательный. Состоит из нервных волокон, отходящих от нейронов ядра, расположенного в ножках мозга на уровне нижних холмиков среднего мозга. Нерв выходит из полости черепа через верхнюю глазничную щель в глазницу и иннервирует верхнюю косую мышцу глазного яблока.
Тройничный нерв ( V пара) – по функции смешанный. Состоит из чувствительных и двигательных нервных волокон.
o Чувствительные нервные волокна являются дендритами нейронов тройничного узла. Эти отростки образуют три ветви:
· глазничную ветвь
· верхнечелюстной нерв
· нижнечелюстной нерв
Аксоны клеток тройничного узла составляют чувствительный корешок тройничного нерва, который идет в мозг к чувствительным ядрам. У тройничного нерва имеется несколько чувствительных ядер, клетки которых находятся преимущественно в мосту, и частично в продолговатом мозге, в верхних шейных сегментах спинного мозга и в ножках головного мозга.
От чувствительных ядер аксоны идут в таламус, а затем в кору больших полушарий (нижний отдел позади центральной извилины).
o Двигательные волокна тройничного нерва – это аксоны двигательного ядра, находящегося в мосту. Эти волокна при выходе из мозга образуют двигательный корешок тройничного нерва, который присоединяется к третьей ветви – нижнечелюстному нерву.
Отводящий нерв ( VI пара) – по функции двигательный. Состоит из аксонов нейронов ядра, расположенного в мосту. Выходит из черепа через верхнюю глазничную щель и иннервирует наружную прямую мышцу глазного яблока.
Лицевой нерв ( VII пара) – по функции смешанный. Содержит:
o двигательные соматические волокна. Аксоны отходят от ядра нерва, находящегося в мосту. Иннервируют мимические мышцы лица, часть мышц шеи.
o секреторные парасимпатические волокна. Иннервируют слюнные железы: околоушные, подъязычные, подчелюстные.
o чувствительные вкусовые волокна. Несут афферентную информацию от сосочков языка.
Секреторные парасимпатические и чувствительные вкусовые волокна входят в состав промежуточного нерва, который также имеет парасимпатические и чувствительные ядра в мосту.
Преддверно-улитковый нерв ( VIII пара) – по функции чувствительный. Состоит из:
· улиткового корешка, идущего от звуковоспринимающего аппарата – спирального органа. Ядра связаны с подкорковыми слуховыми центрами нижних холмиков крыши среднего мозга и медиальных коленчатых тел. Далее волокна идут в слуховую зону коры (средняя часть височной извилины). Ядра нижних холмиков также связаны с ядрами передних рогов спинного мозга, что позволяет осуществлять ориентировочный рефлекс на внезапный звуковой раздражитель.
· преддверного корешка, идущего от вестибулярного аппарата – органа равновесия. Ядра связаны с мозжечком.
Каждая часть преддверно-улиткового нерва имеет свой чувствительный нервный узел.
Языкоглоточный нерв ( IX пара) – по функции смешанный. Содержит:
o чувствительные общие и вкусовые волокна. Несут информацию от слизистой оболочки языка, вкусовых сосочков корня языка, барабанной полости. Имеет два нервных узла.
o двигательные соматические. Иннервируют мышцы глотки.
o секреторные парасимпатические. Иннервируют околоушные слюнные железы.
Все ядра находятся в продолговатом мозге, некоторые из них являются общими с блуждающим нервом.
Блуждающий нерв ( X пара) – по функции смешанный. Содержит:
o чувствительные волокна. Разветвляются в различных внутренних органах, где имеются чувствительные нервные окончания – висцерорецепторы
o двигательные соматические волокна иннервируют поперечнополосатые мышцы глотки, мягкого неба и мышц гортани
o парасимпатические эфферентные волокна иннервируют сердечную мышцу, гладкие мышцы, железы всех внутренних органов грудной полости, живота за исключением сигмовидной кишки и органов малого таза. Имеются два вида парасимпатических волокон:
· двигательные
· секреторные
Ядра чувствительные, двигательные и вегетативные, находятся в продолговатом мозге.
Блуждающий нерв – самый крупный из черепномозговых нервов. В связи с его положением в нем выделяют несколько отделов.
Головной отдел – ветви нерва идут к твердой оболочке мозга, участкам кожи наружного слухового прохода
Шейный отдел – глотка, мягкие мышцы неба, мышцы и слизистая оболочка гортани, верхние сердечные ветви, нерв депрессор – одна из чувствительных ветвей, заканчивающихся в дуге аорты и играющая важную роль в регуляции артериального давления, изменении частоты сердечно-сосудистых сокращений
Грудной отдел – нижние сердечные ветви, ветви к бронхам, к легким, к  пищеводу
пищеводу
Брюшной отдел – нервные сплетения желудка, тонкой кишки, толстой кишки до начала сигмовидной, печень, поджелудочная железа, селезенка, почки, семенники (яичники)
Блуждающий нерв по составу волокон и области иннервации является главным парасимпатическим нервом.
Добавочный нерв ( XI пара) – по функции двигательный. Состоит из аксонов нейронов ядер продолговатого мозга и первого шейного сегмента спинного мозга. Иннервирует грудино-ключично-сосцевидную мышцу и трапециевидную мышцу.
Подъязычный нерв ( XII пара) – по функции двигательный. Ядро находится в продолговатом мозге, иннервирует все мышцы языка, подбородочно-подъязычную мышцу, а также совместно с I – III шейными нервами мышцы шеи, лежащие ниже подбородочно-подъязычной мышцы.
Рилизинг-факторы через капиллярную сеть попадают в гипофиз и способствуют синтезу тропных гормонов. Тропные гормоны гипофиза держат под контролем всю эндокринную систему и обеспечивают гормональный баланс организма.
Гипоталамус играет также центральную роль в реакции организма на стрессовые воздействия.
Задний мозг
11.1. Варолиев мост
11.2. Мозжечок
11.2.1. Общая морфология и проводящие пути мозжечка
11.2.2. Строение коры мозжечка
11.1. Варолиев мост
Варолиев мост (pons Varolii) – это вентральная часть заднего мозга. Представляет собой утолщение в виде поперечного валика, расположенного впереди продолговатого мозга.
В нем различают переднюю часть (основание моста), обращенную к скату черепа и заднюю часть (покрышка моста), обращенную к мозжечку. Граница между ними условно проводится по трапециевидному телу, которое можно обнаружить на фронтальном разрезе моста. Трапециевидное тело состоит из пучков поперечно идущих нервных волокон, относящихся к слуховому проводящему пути.
На передней части моста имеется базиллярная борозда. По бокам моста находятся правая и левая средние ножки мозжечка, в составе которых проходят пучки нервных волокон из моста в мозжечок. Задняя часть моста образует верхнюю половину ромбовидной ямки (IV желудочек).
Мост состоит из:
белого вещества снаружи. Белое вещество моста состоит преимущественно из проводящих путей, проходящих транзитно из других отделов мозга. Все восходящие и нисходящие пути центральной нервной системы, связывающие отделы спинного и головного мозга проходят через Варолиев мост. Передняя часть моста образована в основном белым веществом.
серого вещества внутри. Серое вещество передней части моста представлено собственными ядрами моста. К этим ядрам подходят волокна от коры больших полушарий (коркомостовые пути), а от ядер идут волокна в мозжечок (мостомозжечковые пути). Задняя часть моста содержит большое количество ядер (ядра V, VI, VII, VIII пар черепномозговых нервов). Более того, задняя часть моста является непосредственным продолжением продолговатого мозга.
В Варолиевом мосту локализованы два респираторных центра контролирующие дыхательные функции. Также мост участвует в регуляции вазомоторных реакций, активации коры мозга, обеспечивая, как и ретикулярная формация, реакцию пробуждения. Таким образом, нейроны Варолиева моста являются связующим звеном между продолговатым и средним мозгом, участвуя в регуляциях и интеграциях бульбарного уровня (бульбарный – относящийся к продолговатому мозгу) и среднемозговых влияний на корковые процессы головного мозга.
11.2. Мозжечок
11.2.1. Общая морфология и проводящие пути мозжечка
 Мозжечок (cerebellum), или малый мозг, представляет собой структуру, расположенную над продолговатым мозгом и мостом, позади больших полушарий мозга (рис. 36). Мозжечок состоит из нескольких частей, различных по происхождению в эволюции позвоночных животных.
Мозжечок (cerebellum), или малый мозг, представляет собой структуру, расположенную над продолговатым мозгом и мостом, позади больших полушарий мозга (рис. 36). Мозжечок состоит из нескольких частей, различных по происхождению в эволюции позвоночных животных.
 У человека мозжечок состоит из двух полушарий (hemi - spheria cerebelli), находящихся по бокам от червя (vermis cerebelli). Поверхность мозжечка изрезана глубокими ветвящимися бороздами. Они делят полушария на дольки, которые объединяются в доли:
У человека мозжечок состоит из двух полушарий (hemi - spheria cerebelli), находящихся по бокам от червя (vermis cerebelli). Поверхность мозжечка изрезана глубокими ветвящимися бороздами. Они делят полушария на дольки, которые объединяются в доли:
верхнюю (переднюю)
заднюю
нижнюю (флоккулондулярную)
К филогенетически более древней части мозжечка млекопитающих относят переднюю долю и часть задней доли. Эти структуры мозжечка преимущественно связаны со спинным мозгом и вестибулярным аппаратом, тогда как полушария в основном получают информацию от мышечных и суставных рецепторов, а также от зрительного и слухового анализаторов. Существует классификация, согласно которой кора мозжечка делится на три области (рис. 37):
| |
палеоцеребеллум (древний мозжечок) включает переднюю долю (дольки II - V), простую дольку (долька VI) и заднюю часть корпуса мозжечка (дольки VIII-IX); палеоцеребеллум тесно связан со спинным мозгом, а также имеет двусторонние связи с сенсомоторной областью коры больших полушарий;
неоцеребеллум (новый мозжечок) образован средней частью корпуса мозжечка (долька VII и частично дольки VI и VIII). Сюда поступает информация от коры больших полушарий, а также от слуховых и зрительных рецепторов. Основная часть полушарий мозжечка принадлежит новому мозжечку, лучше всего эта структура развита у человека.
В толще мозжечка находятся три пары ядер (рис. 36, б):
зубчатое, расположенное латерально
ядро шатра – медиально (в черве)
пробковидное и округлое ядра - между ними.
Волокна мозжечка собраны в три пары массивных волокнистых пучков, известных как мозжечковые ножки:
верхние. Через верхние ножки (из среднего мозга) в мозжечок поступают
§ спиномозжечковые пути Говерса
§ волокна от передних бугров четверохолмия (мозжечково-красноядерный путь).
средние. Через средние ножки (из моста) к мозжечку поступают волокна, несущие импульсы от коры больших полушарий (в первую очередь от перецентральной извилины).
нижние. Через нижние ножки (из продолговатого мозга) проходят
§ спинномозговые пути Флексинга
§ волокна от ядер Голля и Бурдаха
§ волокна, идущие от вестибулярного ядра. Все перечисленные волокна несут импульсы от проприо- и вестибулорецепторов.
Как видно из приведенного выше, афферентные волокна входят в мозжечок в основном через нижние и средние ножки мозжечка.
Эфферентные волокна проходят преимущественно через верхние ножки. Они берут начало из центральных ядер и заканчиваются (с перекрестом) в
§ красном ядре. Импульсы, подходящие к красному ядру, через нейроны руброспинального тракта идут к спинному мозгу
§ таламических ядрах. Импульсы, идущие в таламус, далее по таламокорковым путям следуют к моторной зоне коры больших полушарий.
§ стволовой ретикулярной формации
§ подкорковых ядрах
§ продолговатом мозге.
 Однако имеются исключения: некоторая часть спиномозжечковых путей входит через верхние ножки, а некоторые эфферентные волокна от флоккулонодулярной доли и ядра шатра проходят через нижние ножки.
Однако имеются исключения: некоторая часть спиномозжечковых путей входит через верхние ножки, а некоторые эфферентные волокна от флоккулонодулярной доли и ядра шатра проходят через нижние ножки.
Ядра шатра направляют волокна через нижние ножки к вестибулярным ядрам и ретикулярной формации продолговатого мозга и моста. Промежуточные и зубчатые ядра посылают волокна через верхние ножки преимущественно к среднему мозгу и таламусу, в частности, к красному ядру. Основная часть церебелло-таламических волокон отходит от зубчатых ядер. Эти волокна проецируются также к красному ядру, полосатому телу. Подводя итог сказанному, подчеркнем, что влияния мозжечка на спинальные мотонейроны осуществляются через вестибулоспинальные и ретикулоспинальные пути, а на прецентральную область коры - через вентролатеральное ядро таламуса.
Строение коры мозжечка
Кора мозжечка различных представителей позвоночных, включая человека, построена по единому плану и состоит из трех слоев (см. рис 38).
Поверхностный, или молекулярный, слой содержит разветвления дендритов клеток Пуркинье и параллельные волокна. Здесь также расположены звездчатые и корзинчатые клетки
Средний, или слой клеток Пуркинье - содержит тела клеток Пуркинье.
Внутренний зернистый, или гранулярный. Самый нижний, гранулярный слой содержит клетки-зерна, или гранулярные клетки. От тела клетки-зерна отходит 4-7 коротких дендритных отростков, с которыми мшистые волокна образуют синапсы. Аксон клетки-зерна поднимается вертикально вверх и в молекулярном слое Т-образно ветвится, образуя параллельные волокна. Плоскость его ветвления перпендикулярна плоскости ветвления дендритов клеток Пуркинье. В гранулярном слое находятся клетки Гольджи, аксоны которых восходят в молекулярный слой.
Афферентный вход в кору мозжечка осуществляется в основном через две системы волокон:
лазающие. Каждая клетка Пуркинье получает вход только от одного лазающего волокна (медиатор - аспартат), которое заканчивается на начальной (проксимальной) части дендрита. Лазающие волокна представляют собой аксоны нейронов, тела которых находятся в нижних оливах. По этому входу оказывается мощное возбуждающее действие.
моховидные, или мшистые. Моховидные волокна характеризуются обширной дивергенцией (одно волокно образует синапсы примерно на 20 клетках-зернах) и оказывают как тормозное, так и возбуждающее действие на клетки Пуркинье. Возбудительные влияния на клетки Пуркинье от моховидных волокон переключаются через клетки-зерна, аксоны которых поднимаются к поверхности коры мозжечка и, разветвляясь в молекулярном слое, образуют параллельные волокна. Последние оканчиваются возбудительными синапсами (медиатор - глутамат) на дистальных участках дендритов клеток Пуркинье.
Благодаря этому строению коры мозжечок может участвовать в оперативном управлении движениями.
Конечный мозг
12.1. Общая характеристика конечного мозга
12.2. Структура базальных ядер
12.3. Кора больших полушарий
12.1. Общая характеристика конечного мозга
Конечный мозг (cerebrum) – это самый главный и самый крупный отдел ЦНС у позвоночных животных, управляющий деятельностью других отделов мозга и спинным мозгом. На рис. 39 представлена схема отношений основных элементов конечного мозга.
 Это самая новая структура ЦНС, наиболее хорошо она развита у высших позвоночных животных – млекопитающих. У низших позвоночных полушария конечного мозга выполняют лишь чувствительную (в основном обонятельную функцию). У высших позвоночных конечный мозг является структурой, предназначенной для выполнения сложных интегративных, аналитических и рефлекторных функций.
Это самая новая структура ЦНС, наиболее хорошо она развита у высших позвоночных животных – млекопитающих. У низших позвоночных полушария конечного мозга выполняют лишь чувствительную (в основном обонятельную функцию). У высших позвоночных конечный мозг является структурой, предназначенной для выполнения сложных интегративных, аналитических и рефлекторных функций.
Полушария конечного мозга (hemispheria cerebri) разделены продольной мозговой щелью. Полушария сильно разрастаются и покрывают собой все остальные отделы головного мозга. Каждое полушарие имеет свою щелевидную полость – боковой желудочек. Условно желудочек левого полушария считается I, а правого – II. Каждый боковой желудочек состоит из центральной части, от которой в лобную долю отходит передний рог, в глубину затылочной доли – задний рог и отходящий от него вниз и латерально – нижний рог (является полостью височной доли). Боковые желудочки сообщаются с полостью III желудочка и имеют в своих стенках сосудистое сплетение. Медиальной стенкой переднего рога является тонкая прозрачная пластинка. Плавая и левая пластинки образуют между передними рогами общую прозрачную перегородку (septum pellucidum).
 Расположение белого и серого вещества в конечном мозге сходно с таковым в мозжечке: в глубине белого вещества расположены ядра серого вещества, а вся поверхность полушарий покрыта серым веществом - корой.
Расположение белого и серого вещества в конечном мозге сходно с таковым в мозжечке: в глубине белого вещества расположены ядра серого вещества, а вся поверхность полушарий покрыта серым веществом - корой.
 |
12.2. Структура базальных ядер
Термин базальные ядра (или, как их еще называют базальные ганглии) означает, что эти ядра расположены у основания больших полушарий, латеральнее промежуточного мозга. Они составляют примерно 3% от объема полушарий. По общей структуре они подобны стволовым ядрам и разбиты на отдельные группы прослойками белого вещества. Из-за этого они имеют несколько пестрый внешний вид, что отразилось в объединяющем их названии – полосатое тело (corpus striatum).
Кроме полосатого тела (рис. 40) в состав базальных ганглиев (рис. 41) также входят ограда - тонкая прослойка серого вещества, расположенная латеральнее чечевицеобразного ядра и отделенная от него перегородкой из белого вещества (наружной капсулой) и миндалевидное тело (миндалина) – расположенная под чечевицеобразным ядром в самом переднем отделе височной доли. К базальным ядрам относят также субталамические ядра и черную субстанцию.
Анатомически хвостатое ядро тесно связано с боковым желудочком.
Расположенная впереди и медиально его расширенная часть – головка ядра образует боковую стенку переднего рога желудочка.
Тело ядра – нижнюю стенку центральной части желудочка
Тонкий хвост ядра – верхнюю стенку нижнего рога.
Скорлупа и хвостатое ядро имеют слоистую структуру. Бледный шар не имеет слоистости и выглядит светлее остального стриатума. Скорлупа образует наружный слой чечевицеобразного ядра, а бледный шар – внутренние его части. Бледный шар, в свою очередь, состоит из наружного и внутреннего члеников.
Друг от друга хвостатое и чечевицеобразное ядра отделены прослойкой белого вещества – частью внутренней капсулы. Другая часть внутренней капсулы отделяет чечевицеобразное ядро от нижележащего таламуса.
Стриатума делят по филогенетическому признаку на:
Палеостриатум – эволюционно более старая часть, образована бледным шаром
Неостриатум – эволюционно молодая часть, образована хвостатым ядром и скорлупой.
Бледный шар состоит из крупных нейронов, напоминающих мотонейроны и мелких – вставочных нейронов. Большие нейроны дают начало быстропроводящим волокнам, которые заканчиваются в красном ядре и черной субстанции среднего мозга, ядрах промежуточного мозга и образуют основной эфферентный выход полосатого тела.
Хвостатое ядро и ограда состоят из мелких клеток и их аксоны направлены, в основном, к бледному шару.
Система этих ядер:
o участвует в образовании экстрапирамидной системы и связана с регуляцией сложных движений и автоматических движений.
o задействована в переработке сенсорной информации, под ее контролем находятся центры гипоталамуса.
o является высшим регуляторным центром вегетативных функций терморегуляции и углеводного обмена.
Хвостатое ядро участвует в формировании условных рефлексов, механизмов памяти, эмоциях.
Стриатум не получает прямых входов от спинного мозга и от сенсорных ядер мозгового ствола. Основные входы эта структура получает от:
коры больших полушарий. Все корковые проекции на стриатум организованы ипсилатерально (на одноименную сторону тела), за исключением полей 4, 6 и 8, которые образуют билатеральные проекции (на обе стороны тела)
от некоторых ядер таламуса
от черной субстанции
от миндалевидного тела.
Бледный шар и черная субстанция являются основными эфферентными структурами стриарной системы. Эти структуры отсылают волокна к таламусу, в область среднего мозга и каудально в область моста. Их синаптические окончания оказывают тормозное синаптическое действие (медиатор – g - аминомасляная кислота).
Синдром поражения стриарной системы в клинике описывается как атетоз и хорея. При атетозе на фоне повышенного тонуса мышц наблюдаются медленные непрекращающиеся движения, в которые обычно вовлекаются руки, реже - губы и язык, а иногда - шея и ноги. При хорее тонус мышц также повышен, но движения быстрые, отрывистые. Они нерегулярны и перемежаются в непредсказуемой последовательности.
Хорея является ведущим моторным симптомом и при болезни Гетингтона, которая характеризуется разрушением эфферентных нейронов стриарной системы. При двустороннем поражении субталамического ядра (тела Люиса) в непроизвольную моторную активность вовлекается дистальная мускулатура конечностей. Это так называемый баллизм. При повреждении субталамического ядра только одной стороны непроизвольные движения наблюдаются лишь на противоположной стороне тела (гемибаллизм).
Изолированное повреждение эфферентных нейронов внутреннего сегмента бледного шара и ретикулярной части черной субстанции у экспериментальных животных не вызывает заметных двигательных расстройств, за исключением того, что все движения замедляются (брадикинезия).
У человека система связей между стриарной системой и черной субстанцией (ведущий медиатор – дофамин) наиболее обширно повреждается при болезни Паркинсона. Основными моторными нарушениями в этом случае являются брадикинезия, ригидность (оцепенелость, обусловленная напряжением мышц) и тремор покоя (дрожание, ритмичные колебательные движения конечностей или головы; тремор покоя уменьшается при движении, но не исчезает под действием алкоголя). Эти симптомы в полном объеме воспроизводятся у обезьян при экспериментальном разрушении дофаминовых нейронов мозга. В основе болезни Паркинсона лежит разрушение нейронов, приводящее к уменьшению содержания медиатора дофамина в стриатуме и нарушения баланса дофаминергических и холинергических влияний.
Кора больших полушарий
В каждом полушарии выделяют пять долей: лобную, теменную, затылочную, височную и островковую (островок). Эти доли отделены друг от друга глубокими бороздами. Центральная борозда (Роландова) отделяет лобную долю от теменной, латеральная борозда (Сильвиева) – височную от лобной и теменной, темнено–затылочная борозда разделяет теменную и затылочную доли, а в глубине латеральной борозды располагается островковая доля. Борозды и извилины больших полушарий образуют так называемый плащ и значительно увеличивает его площадь.
Каждое полушарие покрыто тонкой пластинкой серого вещества называемого корой. Всю кору делят на:
древнюю
старую
новую.
К древней и старой коре относятся те области, которые преимущественно связаны с регуляцией вегетативных функций, эмоций, инстинктов и некоторых видов памяти.
Новая кора достаточным образом развита только у млекопитающих. Степень ее развития оценивается коэффициентом цефализации: отношение массы мозга к массе тела.
Только у летучих мышей этот коэффициент выше, чем у человека. Это связано с облегчением их тела. Еще одним из объективных признаков прогрессивного развития новой коры является увеличение общего количества нейронов и их связей. В настоящее время считается, что число нейронов в коре человека достигает 12-18 млрд., а ее толщина в разных участках мозга колеблется от 1,3 до 5 мм, площадь же коры составляет в среднем 2500см2.
|
В коре выделяют шесть слоев, которые состоят из нервных и глиальных элементов (рис. 42).
Нервные клетки по своей форме весьма разнообразны. Среди них выделяют 56 разновидностей, но более обобщенно выделяют 3 вида:
пирамидные. Среди пирамидных клеток различают большие и малые. Большие пирамидные клетки – это эфферентные (двигательные)
звездчатые
веретеновидные.
 1) Первый наружный слой называется молекулярным. Он состоит из окончаний дендритных отростков и мелких мультиполярных ассоциативных нейронов.
1) Первый наружный слой называется молекулярным. Он состоит из окончаний дендритных отростков и мелких мультиполярных ассоциативных нейронов.
2) Второй слой – наружный зернистый, состоит из вставочных нейронов.
3) Третий слой – пирамидный, состоит из мелких разной величины пирамидных клеток.
4) Четвертый - внутренний зернистый, также образованный вставочными нейронами.
5) В пятом слое – ганглиозном (внутреннем пирамидном) располагаются большие пирамидные клетки, которые описал Бец и поэтому они еще называются клетками Беца.
6) Шестой слой коры называется полиморфным и состоит из звездчатых и веретеновидных нейронов.
Толщина слоев, характер их границ, размеры клеток и их количество варьируют в разных отделах головного мозга. Колонки в пределах каждой зоны и между зонами объединяются в морфо–функциональные единицы, реагирующие на определенный характер раздражения в данный период времени и формируют распределительные системы.
По функциональному признаку нейроны коры подразделяются на:
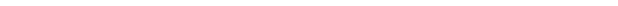 сенсорные. К ним подходят волокна восходящих (афферентных) проводящих путей.
сенсорные. К ним подходят волокна восходящих (афферентных) проводящих путей.
промежуточные. Осуществляют связь с разными участками коры.
эффекторные. Нервные клетки, отростки которых образуют нисходящие (эфферентные) проводящие пути, идущие к различным ядрам головного и спинного мозга.
Цитоархитектонические особенности (состав клеток коры, толщина слоев) позволили разделить всю поверхность коры на 10 цитоархитектонических областей, включающих в себя 52 поля – поля Бродмана (по фамилии немецкого физиолога).
Бродман разделил кору каждого полушария на следующие области: затылочную, верхнюю теменную, нижнюю теменную, постцентральную, центральную, прецентральную, лобную, височную, лимбическую, островковую.
Каждое цитоархитектоническое поле обозначено на картах номером, который присваивался ему в порядке описания. Следует отметить, что между полями не существует резких границ, клеточные слои плавно меняют свою структуру при переходе от данного поля к другому (рис. 43).
Павлов рассматривал кору как совокупность корковых (или мозговых) концов анализаторов (зрительного, слухового, обонятельного и др.). В них происходит анализ информации, поступившей из органов чувств (рецепторов) по восходящим (афферентным) проводящим путям. Каждый корковый конец анализатора состоит из центральной части (или ядра) и периферической части (или «рассеянных элементов»). В клинике вместо термина «корковый конец анализатора» употребляют термин «зона». При этом выделяют:
сенсорные (чувствительные)
двигательные (моторные) зоны коры.
Основные зоны коры:
1-я зона – двигательная – представлена центральной извилиной и лобной зоной впереди нее – 4, 6, 8, 9-е поля Бродмана. От начала больших пирамидных клеток (слой 3) прецентральной извилины берет начало пирамидный нисходящий путь (путь произвольных движений).
2- я зона – чувствительная – участки коры позади центральной борозды (1, 2, 3, 4, 5, 7-е поля Бродмана). Зона кожной чувствительности находится в постцентральной извилине и в заднем отделе парацентральной дольки. В эту зону через ядра таламуса поступает информация от рецепторов кожи противоположной стороны тела. Кора постцентральной извилины является зоной мышечно-суставной чувствительности (информация от проприорецепторов).
3-я зона – зрительная – расположена на медиальной поверхности затылочной доли по краям шпорной борозды (поля 17, 18, 19). Сюда через латеральные коленчатые тела и бугры четверохолмия (подкорковые центры зрения) поступает информация от рецепторов сетчатки глаза. Зона каждого полушария связана с сетчаткой обоих глаз
4-я зона – слуховая – височная область коры головного мозга (22, 41, 42-е поля). В зону поступает информация из рецепторов спирального органа улитки через медиальные коленчатые тела и нижние бугры четверохолмия.
5-я зона – обонятельная – располагается преимущественно в парагиппокампальной извилине на височной доле (11-е поле)
6-я зона – вкусовая – 43-е поле Бродмана.
7-я зона – речедвигательная (по Джексону – центр речи) – у большинства людей (праворуких) располагается в левом полушарии. Состоит из трех отделов:
o Речедвигательный центр Брока (поля 44, 45) - двигательный центр мышц языка. При повреждении этой зоны развивается моторная афазия (а – отрицание, phasis –речь).
o Сенсорный центр Вернике (поля 41, 42) – в заднем отделе височной извилины – связан с восприятием устной речи.
o Центр восприятия письменной речи в зрительной зоне коры головного мозга (18-е поле Бродмана)
В коре головного мозга существуют три проекционные зоны:
Первичная проекционная зона занимает центральную часть ядра мозгового анализатора. Это совокупность наиболее дифференцированных нейронов, в которых происходит высший анализ и синтез информации, возникают четкие и сложные ощущения. К этим нейронам подходят импульсы по специфическому пути передачи импульсов в кору головного мозга (спиноталамический путь)
Вторичная расположена вокруг первичной, входит в состав мозгового отдела анализатора. Обеспечивает сложное восприятие. При поражении этой зоны возникает сложное нарушении функции. Чувствительность к раздражителям при этом обычно не нарушается, но нарушается способность к интерпретации значения раздражителя.
Третичная проекционная зона – ассоциативная – это полимодальные нейроны, разбросанные по всей коре головного мозга. К ним поступают импульсы от ассоциативных ядер таламуса и конвергируют импульсы различной модальности. Обеспечивает связь между различными анализаторами и участвует в формировании условных рефлексов.
Чтобы перейти к раскрытию вопроса об организации лимбической системы необходимо сказать, что кора больших полушарий с филогенетической точки зрения подразделяется на:
Археокортекс – древняя кора. Включает в себя обонятельные луковицы, в которые поступают обонятельные волокна, идущие от эпителия носа, обонятельные тракты и обонятельные бугорки.
Палеокортекс – старая кора. Состоит из поясной извилины, извилины морского конька (гиппокамп) и миндалевидного тела (миндалина, амигдала).
Неокортекс – все остальные обрасти коры.
Раздел 5
ВЕГЕТАТИВНАЯ РЕГУЛЯЦИЯ
Раздел 6
ЭНДОКРИННАЯ РЕГУЛЯЦИЯ
Эндокринная система
15.1. Общие данные о строении гипофиза
15.2. Единая нейрогуморальная регуляция организма
Эндокринная система (от греч. endo – внутрь, krino – выделяю, синоним - железы внутренней секреции) наряду с нервной системой занимает одно из центральных мест в управлении различными процессами жизнедеятельности на уровне целого организма. Центральный отдел эндокринной системы, также как и центральный отдел нервной системы находится внутри черепа. Это придаток мозга, известный под названием – гипофиз (hypophysis). Он представляет собой небольшую железу овальной формы, находящуюся в основании мозга. Сверху гипофиз прикрыт отростком твердой мозговой оболочки.
С помощью, так называемой, гипофизарной ножки гипофиз связан с воронкой, которая отходит от серого бугра гипоталамуса. Центральный отдел эндокринной системы оказывается тесно связанным с центральным отделом нервной системы. Эта связь обеспечивается двумя основными механизмами:
при помощи отростков нервных клеток: их синаптических контактов и нейроэндокринных влияний
при помощи взаимодействий через общее кровеносное русло: передняя гипофизарная артерия образует сеть, общую для гипоталамуса и передней доли гипофиза.
Периферически расположенные железы внутренней секреции также тесно связаны с нервной системой, преимущественно с вегетативной, и снабжены большим количеством кровеносных сосудов. Последнее определяется не только тем, что секретируемые гормоны выделяются непосредственно в кровь, но и с высоким уровнем обменных процессов в этих железах.
Словарь терминов
Автономная нервная система – (вегетативная нервная система) часть нервной системы, иннервирующая висцеральные органы, кожу, гладкие мышцы, железы и сердце; состоит из симпатического и парасимпатического отделов.
Адреналин - гормон, выделяемый мозговым слоем надпочечников; его действие в некоторых отношениях аналогично влиянию симпатических нервов.
Адренергический - относится к нейронам, продуцирующим норадреналин (адреналин) как медиатор.
Аксон - отросток нейрона, по которому возбуждение (потенциал действия) проводится от данного нейрона к другой клетке (другому нейрону, мышечной или железистой клетке).
Аксональный транспорт - движение веществ вдоль аксона.
Аксонный холмик - область тела нейрона, от которой берет начало аксон; обычно является местом генерации импульсов.
Аксоплазма - внутриклеточная жидкая среда аксона.
Активная зона - область пресинаптической терминали, характеризующаяся плотными скоплениями на цитоплазматической поверхности пресинаптической мембраны и скоплениями синаптических пузырьков; считается местом выброса медиатора.
Активная зона синапса - специализированные места цитоплазматической мембраны пресинаптической терминали, в которых происходит освобождение медиатора, упакованного в пузырьки.
Амигдала см. Миндалина.
Ассоциативные области коры - в коре больших полушарий выделяют три ассоциативные зоны: теменно-височную, префронтальную и лимбическую.
Астроцит - один из видов глиальных клеток ЦНС позвоночных.
Афферент - нейрон, или путь, который передает сигналы к центральной нервной системе. Например, слуховые афференты, вестибулярные афференты и т.д.
Афферентные аксоны - аксоны, проводящие импульсы по направлению к центральной нервной системе.
Ацетилхолин - медиатор, выделяющийся в нервно-мышечных синапсах и некоторых центральных синапсах.
Базальные ганглии, или Стриатум - ядра больших полушарий мозга. Включают бледный шар, хвостатое ядро и скорлупу. Проводящими путями тесно связаны с черной субстанцией, субталамическим ядром (телом Люиса).
Белое вещество - часть центральной нервной ткани, состоящая из миелинизированных нервных волокон.
Биогенные амины - общее название биологически активных аминов, включающее норадреналин, адреналин, дофамин, серотонин и гистамин.
Биполярная клетка - нейрон, обладающий двумя крупными отростками, отходящими от тела клетки; в сетчатке позвоночных биполяры находятся между слоями рецепторных и ганглиозных клеток.
Большие полушария - парные структуры головного мозга, особенно хорошо развитые у человека и высших обезьян, связаны между собой мозолистым телом.
Брока зона - область лобной коры, критически задействованная в экспрессивной речи.
Вегетативная нервная система - часть нервной системы, иннервирующая внутренние органы, кожу, гладкую мускулатуру, железы и сердце; в ней различают два отдела: парасимпатическую и симпатическую нервную систему.
Вентральные корешки спинного мозга - образованы аксонами мотонейронов передних рогов серого вещества спинного мозга, а также аксонами нейронов (симпатических) боковых рогов серого вещества спинного мозга грудных сегментов.
Вернике зона - область коры на стыке височной и теменной долей, критически задействованной в сенсорном анализе речи.
Возбуждение - процесс, ведущий к генерации потенциала действия (нервного импульса).
Волокно нервное - аксон нейрона, который находится в составе периферического нерва.
Вставочный нейрон - нейрон, который не является ни сенсорным, ни двигательным, а осуществляет связь между другими нейронами.
Ганглий - обособленное скопление нервных клеток, находящееся (у позвоночных) вне центральной нервной системы.
Гематоэнцефалический барьер - термин, означающий ограничение доступа веществ к нейронам и глиальным клеткам внутри мозга.
Гипоталамус - нижняя часть промежуточного мозга, составляет стенки III желудочка мозга. Входит в состав лимбической системы мозга. Является главной структурой мозга, обеспечивающей биологические мотивации (потребление пищи, половое поведение, поддержание водно-солевого баланса организма и многое другое).
Гипофиз - нижняя мозговая железа, один из основных эндокринных органов мозга. Имеет в своем составе три доли: переднюю, или аденогипофиз, заднюю, или нейрогипофиз, и промежуточную. Гипофиз связан с гипоталамусом гипоталамо-гипофизарным трактом.
Гиппокамп - структура старой коры на медиальной стороне височных долей. Повреждение гиппокампа приводит к синдрому Корсакова. Он тесно связан с гипоталамусом волокнами свода (fornix).
Глия - клетки, входящие в состав нервной ткани. Обычно выделяют астроциты (преимущественно находятся в сером веществе мозга), олигодендроциты (преимущественно находятся в белом веществе мозга) и микроглию, участвующую в образовании мозговых оболочек.
Гормон - биологически активное вещество, выделяющееся из желез внутренней секреции.
Дендрит - от греч. «дерево». Разветвленный отросток нейрона, на котором оканчиваются синапсами многочисленные аксоны других нейронов.
Дорсальные корешки спинного мозга - составлены центральными отростками псевдоуниполярных нейронов межпозвоночных ганглиев. Занимают задний (дорсальный) канатик спинного мозга.
Желудочки - полости в мозгу, заполненные спинномозговой жидкостью; стенки желудочков выстланы эпендимальными клетками.
Зрительный перекрест (хиазма) - место перекрещивания части волокон зрительных нервов. У кошки и у приматов волокна от медиальной области сетчатки идут к латеральному коленчатому телу противоположной стороны.
Интрафузальное волокно - мышечное волокно, входящее в состав веретена; его сокращение приводит к генерации или модуляции сенсорного разряда.
Ипсилатеральный - относящийся к той же стороне тела.
Катехоламины - общее название молекул, имеющих катехольное кольцо и аминогруппы; обычно имеются в виду дофамин, норадреналин и адреналин.
Каудальный - задний (у животных).
Колонки коры - группы корковых нейронов, обладающих одинаковыми свойствами (например, в отношении сенсорной модальности, положения или ориентации рецептивного поля и т.п.).
Комиссура - группа аксонов, соединяющих отдельные структуры мозга. Например, самая большая комиссура - мозолистое тело, которое соединяет большие полушария.
Контралатеральный - относящийся к противоположной стороне тела.
Концевая пластинка - постсинаптическая область скелетного мышечного волокна у позвоночных.
Кора больших полушарий, или Плащ (palium) - покрывает большие полушария преимущественно (95%) новой корой (имеет в своем составе 6 слоев), но также старой (трехслойная кора) - около 2,5% и древней корой (слои нейронов выражены неясно).
Латеральное коленчатое тело — ядро, которое находится в задне-нижней части таламуса и осуществляет функцию реле в зрительном пути.
Лимбическая система - группа структур мозга, связанных между собой тесными связями. Участвует в обеспечении мотивационно-эмоциональной деятельности организма. Одной из главных структур лимбической системы является гипоталамус, которым большинство структур объединены в целостную систему, регулирующую мотивационно -эмоциональные реакции человека и животных на внешние стимулы.
Медиатор - вещество, которое освобождается из нервных терминалей и воздействует на мембрану постсинаптической клетки, обычно вызывая повышение проницаемости мембраны для определенных ионов.
Миелиновая оболочка - обладающая высоким сопротивлением оболочка вокруг аксона, образованная слившимися мембранами шванновских или глиальных клеток.
Миндалина - группа ядер, локализованных в глубине переднего полюса височной доли мозга. Имеет тесные связи с гипоталамусом, гиппокампом, таламусом, с обонятельной системой. Является частью лимбической системы мозга. Координирует эндокринные ответы и реакции автономной нервной вегетативной системы, связанные с эмоциями.
Модальность - близкая группа ощущений. Например, зрительная модальность объединяет ощущение света, темноты, цвета и другие характеристики зрительного стимула. Термин «модальность» часто употребляют для обозначения стимула; например, слуховая модальность - стимулы, адресованные слуховому анализатору.
Мозговой ствол - включает продолговатый мозг, мост и средний мозг. Содержит в своем составе ядра черепных нервов, ретикулярную формацию.
Мозжечок - (лат. Cerebellum - малый мозг) - находится над мостом; состоит из двух полушарий и червя между ними, в глубине полушарий находится четыре пары ядер (ядра шатра, округлое, пробковидное и зубчатое). Мозжечок участвует в управлении движениями.
Мозолистое тело - многочисленные нервные волокна, соединяющие между собой симметричные точки коры больших полушарий.
Мотонейрон (двигательный нейрон) - нервная клетка, аксон которой иннервирует мышечные волокна.
Мышечное веретено - концевой орган в составе скелетной мышцы, от которого берут начало афферентные сенсорные волокна и на котором образуют окончания несколько мотонейронов. Является собственным (проприорецептором) рецептором мышцы. Состоит из группы очень тонких мышечных волокон (интрафузальных волокон), собранных в структуру, напоминающую веретено. Мышечное веретено получает два типа волокон: сенсорные волокна, по которым информация о состоянии веретена передается в спинной мозг, и моторные волокна (аксоны гамма-мотонейронов), возбуждение по которым управляет длиной мышечного веретена.
Начальный сегмент - участок аксона, примыкающий к телу клетки; часто является местом генерации нервных импульсов.
Нейроглия - совокупность клеток-сателлитов, ассоциированных с нейронами. В центральной нервной системе млекопитающих нейроглия образована двумя основными видами клеток — астроцитами и олигодендроцитами; в периферических нервах клетки-сателлиты называются шванновскими клетками.
Нейрон - главная клетка нервной ткани. Состоит из тела (сомы) и двух отростков: многочисленных дендритов и одного аксона. Способен возбуждаться и по аксону проводить возбуждение к другим клеткам организма.
Нейропиль - сплетение тонких аксонов и дендритов; содержит большое число синапсов.
Норадреналин - медиатор, освобождаемый из большинства симпатических нервных терминалей.
Перехват Ранвье - регулярные (для периферических волокон в среднем через 1,5 мм) места аксона, не покрытые миелином. Это связано с тем, что миелиновый чехол образуется оборачиванием вокруг аксона отдельных глиальных клеток. На стыке между отдельными клетками и образуется перехват Ранвье.
Поля 17, 18, 19 — зрительные области коры больших полушарий; различаются по определенным гистологическим и физиологическим критериям.
Ретикулярная формация - (от лат. reticulo - сетчатый) - скопление диффузно расположенных нейронов в сердцевине мозгового ствола и медиальных частях таламуса.
Рефлекс - непроизвольное движение или иная периферическая реакция, вызываемая периферическим стимулом и опосредуемая через центральную нервную систему.
Рецептивное поле — периферическая область, раздражение которой оказывает влияние на разряд данного нейрона. Для нейронов зрительного пути рецептивное поле представляет собой область сетчатки, освещение которой сказывается на активности нейрона.
Рецептор - 1) сенсорное нервное окончание; 2) молекула в составе клеточной мембраны, вступающая в соединение со специфическим веществом - медиатором.
Сагиттальный разрез - вертикальный разрез в передне-заднем направлении.
Серое вещество - часть центральной нервной ткани, образованная преимущественно телами нервных клеток и тонкими нервными окончаниями (в отличие от белого вещества, состоящего из крупных нервных путей).
Серотонин - 5-окситриптамин, служит медиатором в нервной системе моллюсков, а возможно, и в центральной нервной системе позвоночных.
Синапс - контакт аксона с мембраной другой клетки (нейрона, мышечной, железистой и др.), термин был предложен Шеррингтоном.
Синаптическая щель - пространство между мембранами пре- и постсинаптической клетки, через которое должен диффундировать медиатор (в химическом синапсе).
Синаптические пузырьки - небольшие, окруженные мембраной пузырьки, находящиеся в нервных терминалях. Пузырьки с электронноплотной сердцевиной содержат катехоламины и серотонин; прозрачные пузырьки считаются хранилищами других медиаторов, освобождаемых из терминалей.
Сосудистое сплетение - богатое кровеносными сосудами ворсинчатое образование, которое выдается в полость мозгового желудочка и секретирует спинномозговую жидкость.
Спинной мозг - самая каудальная (задняя) часть центральной нервной системы. Имеет сегментарное строение. Обеспечивает двустороннюю связь туловища и ЦНС; способен к автономной деятельности по рефлекторному принципу, например способен осуществлять многие двигательные (коленный, сухожильный и ряд других) и вегетативные (просвет периферических кровеносных сосудов, потоотделение и ряд других) рефлексы.
Спинномозговая жидкость - прозрачная жидкость, заполняющая мозговые желудочки и пространства между твердой, паутинной и мягкой оболочками.
Стриарная кора - называется также полем 17 или зрительным полем 1 - первичная зрительная область затылочной доли мозга, различаемая невооруженным глазом по полосам Геннари.
Субарахноидальное пространство - заполненное спинномозговой жидкостью пространство между двумя слоями соединительной ткани - оболочками, окружающими мозг.
Таламус - дорсальная (верхняя) часть промежуточного мозга. Содержит ядра (скопления нейронов), в которых возбуждение переключается на кору. Образно говоря, таламус является коллектором (собирателем и распределителем) импульсации, поступающей к коре больших полушарий.
Томография - неинвазивный (без проникновения) метод исследования структур головного мозга. Наиболее распространены рентгеновский, позитрон-эмиссионный и магнитно-резонансный методы томографии.
Торможение - влияние одного нейрона на другой, затрудняющее генерацию импульсов.
Фронтальный разрез - разрез, проходящий через череп под прямым углом к продольной (сагиттальной) оси.
Холинэргические нейроны - нейроны, освобождающие в качестве медиатора ацетилхолин.
Центральная нервная система - (ЦНС) - включает в свой состав спинной и головной мозг. Противопоставляется нервной периферической системе.
Центральный нейрон - нейрон ЦНС.
Шванновская клетка - клетка-сателлит в периферической нервной системе; участвует в формировании миелиновой оболочки.
Щелевой контакт - специализированный контакт между клетками - место электрической связи, пространство между прилегающими мембранами невелико - до 2 нм.
Экстрафузальные мышечные волокна - волокна, составляющие основную массу скелетной мышцы (кроме мышечных веретен).
Электроэнцефалограмма - запись электрической активности мозга при помощи внешних электродов, приложенных к черепу.
Эндокринный орган - (железа), или Железа внутренней секреции - выделяет секреты в среду организма; специальные протоки отсутствуют.
Эндотелиальные клетки, эндотелий - клетки, выстилающие кровеносные сосуды.
Эпендима - слой клеток, выстилающих мозговые желудочки и центральный канал спинного мозга.
Эфферентные аксоны - нервные волокна, проводящие импульсы из центральной нервной системы на периферию.
g -Аминомасляная кислота (ГАМК) - тормозной медиатор в нервно-мышечных синапсах ракообразных и, возможно, в центральной нервной системе позвоночных.
Литература
Основная
Воронова Н.В., Климова Н.М., Менджерицкий А.М. Анатомия центральной нервной системы. М.: Аспект пресс, 2005.
Гаврилов Л.Ф., Татаринов В.Г. Анатомия. М.: Медицина, 1978.
Сапин М.П., Билич Г.Л. Анатомия человека: В 2 кн. 2-е изд., перераб. и доп. М. 1999.
Шульговский В. В. Основы нейрофизиологии: Учебное пособие для студентов вузов. - М.: Аспект Пресс, 2000. с. 277.
Дополнительная
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Роберте К., Уотсон Дж. Молекулярная биология клетки: В 3-х т. 2-е изд., Пер. с англ.-М.: Мир, 1994.
Богданов А.В. Физиология центральной нервной системы. М.: Изд-во УРАО, 2002.
Моренков Э. Д. Морфология мозга человека. М.: Изд-во Моск. ун-та, 1978.
Немечек С., Лодин З., Вольф И.Р., Выскочил Ф., Байгар. И. Введение в нейробиологию. ЧССР Прага: Мед. изд-во Авиценум. Изд-е 1-ое русское – 1978.
Николс Дж., Мартин Р., Валлас Б., Фукс П. От нейрона к мозгу / Пер. с англ. П. М. Балабана, А.В.Галкина, Р. А. Гиниатуллина, Р.Н.Хазипова, Л.С.Хируга. — М.: Едиториал УРСС, 2003
Корнев М.А., Кульбах О.С. Основы строения центральной нервной системы. С-Пб.: Фолант, 2002
Коновалов А. Н., Блинков С. М., Пуцило М. В. Атлас нейрохирургической анатомии. М., 1990.
Оленев С. Н. Развивающийся мозг. Л., 1979.
Савельев С. Д. Стереоскопический атлас мозга человека. М.: Area XVII, 1996.
Сапелкин А. А. Анатомия человека. Владивосток. Изд-во ДВГУ, 2001.
Шаде Дж., Форд П. Основы неврологии. М., 1976.
Хэм А., Кормак Д. Гистология. Т.3. М.: Мир, 1983.
Содержание
Предисловие ………………………………………………….............................4
1. Введение ……………………………………………………………………...5
1.1. Содержание предмета «Анатомия ЦНС»……………………..……5
1.2. Методы……………………………………………..…………………6
1.3. Задачи …………………………………………………..…………….6
1.4. Строение нервной системы ………………………………………….7
1.5. Анатомическая номенклатура……………………………………….9
Раздел 1
НЕРВНАЯ ТКАНЬ
2. Нейроны – специализированные возбудимые клетки нервной системы…………………………………………………………………………11
2.1. Структура нейрона………………………………………….………11
2.2. Ультраструктурные особенности нервных клеток………………...11
2.2.1. Клеточная мембрана……………………………………….....14
2.2.2. Цитоплазма и клеточные органеллы………………………...16
2.3. Классификация нейронов………………………………...…………18
3. Синапсы…………………………………………………………………...…20
3.1. Общая характеристика синапсов………………………………….20
3.2. Электрический синапс …………………………………………….22
3.3. Химический синапс………………………………………………..23
3.4. Рефлекторная дуга……………………………………………………25
4. Глия. Строение нервов……………………………………………….......27
4.1. Общая характеристика клеток глии………………………………27
4.2. Типы глиальных клеток ……………………………………………..28
4.2.1. Эпендима……………………………………………………...29
4.2.2. Астроцитарная глия………………………………………….30
4.2.3. Олигодендроглия……………………………………………..31
4.2.4. Микроглия…………………………………………………………..31
4.3. Строение нервов……………………………………………………...32
Раздел 2
РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ
5. Развитие нервной системы в филогенезе………………………..34
5.1. Диффузный тип нервной системы………………………..…34
5.2. Ганглионарный тип нервной системы……………………………37
5.3. Трубчатый тип нервной системы…………………………………40
6. Развитие нервной системы в онтогенезе…………………………...…42
6.1. Индивидуальное развитие организма…………………………….42
6.2. Онтогенез нервной ткани………………………………………….43
6.3. Детерминация и дифференцировка нейронов в онтогенезе…….47
Раздел 3
СПИННОЙ МОЗГ
7. Общая морфология спинного мозга………………………………………50
7.1. Общая характеристика спинного мозга……………………………50
7.2. Внутренне строение спинного мозга………………………………54
7.3. Мозговые оболочки…………………………………………………55
8. Организация серого и белого вещества спинного мозга……………..57
8.1. Строение серого вещества спинного мозга………………………..57
8.2. Основные проводящие пути спинного мозга ……………………..58
Раздел 4
ГОЛОВНОЙ МОЗГ
9. Обзор головного мозга………………………………………………….62
9.1. Общая характеристика головного мозга…………………………62
9.2. Основание мозга……………………………………………………...64
9.3. Черепномозговые нервы……………………………………………..66
9.4. Сагиттальный срез головного мозга………………………………..71
10. Анатомия ствола головного мозга ……………………………………72
10.1. Продолговатый мозг……………………………………………….73
10.2. Средний мозг ………………………………………………………74
10.3. Промежуточный мозг……………………………………………...75
10.4. Ретикулярная формация ствола мозга……………………………79
11. Задний мозг………………………………………………………………….80
11.1. Варолиев мост……………………………………………………...80
11.2. Мозжечок…………………………………………………………...80
11.2.1. Общая морфология и проводящие пути мозжечка……....80
11.2.2. Строение коры мозжечка………………………………….85
12. Конечный мозг……………………………………………………………86
12.1. Общая характеристика конечного мозга………………………….86
12.2. Структура базальных ядер………………………………………....87
12.3. Кора больших полушарий……………………………………….....90
13. Лимбическая система и проводящие пути конечного мозга…….95
13.1. Лимбическая система………………………………………………95
13.2. Проводящие пути конечного мозга………………………………..99
Раздел 5
ВЕГЕТАТИВНАЯ РЕГУЛЯЦИЯ
14. Вегетативная нервная система………………………………………..102
14.1. Общая характеристика ВНС…………………………………...102
14.1.1. Центральный отдел вегетативной нервной системы……103
14.1.2. Периферический отдел вегетативной нервной системы..104
14.2. Сравнительная характеристика СПС и ППС…………………….108
Раздел 6
ЭНДОКРИННАЯ РЕГУЛЯЦИЯ
15. Эндокринная система……………………………………………………110
15.1. Общие данные о строении гипофиза……………………………..110
15.2. Единая нейрогуморальная регуляция организма………………111
Приложения…………………………………………………………………117
Словарь терминов……………………………………………………………123
Литература…………………………………………………………………130
Учебное издание
АНАТОМИЯ ЦНС
Курс лекций
В авторской редакции
Технический редактор Л.И. Гурова
Компьютерная верстка В.А. Рассказовой
Подписано в печать 21.05.2007
Формат 60х84 1/16 Усл. печ. л. ____ Уч-изд. л. ____
Тираж 200 экз. Заказ_________
Издательство Дальневосточного университета
690950, г. Владивосток, ул. Октябрьская, 27
Отпечатано в типографии
Издательско-полиграфического комплекса ДВГУ
690950, г. Владивосток, ул. Алеутская, 56
И. А. Кирсанова
Анатомия ЦНС
курс лекций
для студентов гуманитарных специальностей
Владивосток
2007
Министерство образования и науки Российской Федерации
Федеральное агентство по образованию
Дальневосточный государственный университет
Кафедра клеточной биологии
И. А. Кирсанова
Анатомия ЦНС
курс лекций
для студентов гуманитарных специальностей
Владивосток
Издательство Дальневосточного университета
2007
УДК 612.82(075.8)
ББК 28.706.3
К 43
Ответственный редактор
А.П. Анисимов, докт. биол. наук, профессор
Кирсанова И.А.
Анатомия ЦНС. Курс лекций.-
К 43 Владивосток: Изд–во Дальневост. ун–та, 2007. - с.
В учебном пособии рассматриваются строение и общие принципы организации нейрона и нервной ткани, показаны общие закономерности формирования нервной системы в эволюционном и индивидуальном развитии. Описывается строение и связи отделов центральной нервной системы человека. Особое внимание уделяется структурам мозга, обуславливающим психические функции.
Учебное пособие предназначено для студентов гуманитарных специальностей.
К 2007010000
180 (03) - 2007 ББК 28.706.3
© Кирсанова И.А.
© Издательство Дальневосточного университета, 2007
Предисловие
Анатомия центральной нервной системы (ЦНС) является базовой биологической дисциплиной при подготовке психологов и социальных работников. Фундаментальные данные о строении отделов ЦНС не изменялись в течение многих десятков и даже сотен лет. Вместе с тем активное развитие нейробиологии и нейрофизиологии в последние годы несколько изменило представление об устройстве и функции как нервных клеток, так и отдельных областей нервной системы. В ходе курса студенты получат знания об основных принципах организации нервной системы, познакомятся с закономерностями ее развития в онтогенезе с учетом возрастных особенностей, и в филогенезе – ее историческом, эволюционном развитии.
Для подготовки будущих психологов данная дисциплина является теоретической основой позволяющей понять законы поведения животных, законы становления и функционирования человеческой психики. Последующие дисциплины, усвоение которых опирается на данные «Анатомии ЦНС» - это физиология ЦНС, физиология высшей нервной деятельности и сенсорных систем, психофизиология, нейропсихология, психогенетика и др.
Главной задачей данного курса является формирование у студентов представления о многоуровневой организации нервной системы. К другим, не менее важным задачам относятся:
- создать у студентов представление о ЦНС, как об одной из интегративных систем организма,
- сформировать у студентов представление о нервной системе как материальном субстрате психической деятельности,
- научить студентов распознавать основные структуры мозга, объяснять их значение,
- применять знания об организации и основных принципах работы мозга в смежных дисциплинах.
Подробное описание морфологии ЦНС на субклеточном, клеточном, тканевом и системном уровнях с учетом приведенных иллюстраций облегчит усвоение теоретического материала. Определенную трудность для студентов представляет собой необходимость усвоения терминологии данной дисциплины. В настоящем пособии предпринята попытка представить материал достаточно подробно, с учетом академичности изучаемой дисциплины, в сочетании с его доступностью для студентов-гуманитариев. С той же целью в пособии приведен список научных терминов, которые помогут студентам при самостоятельной работе с использованием фундаментальных учебников по анатомии.
Лекционный курс рассчитан на 37 ч и состоит из шести разделов, включающих 15 тем. В первом разделе « Нервная ткань» рассматривается микроанатомическое строение ЦНС: нейроны и глия, а также строение нервов. Второй раздел «Развитие нервной ткани» содержит информацию о развитии нервной системы в филогенезе и онтогенезе. Третий раздел «Спинной мозг» и четвертый раздел «Головной мозг», посвящены изучению макроанатомии основных отделов ЦНС. Пятый раздел «Вегетативная нервная система» содержит данные об особенностях строения автономной нервной системы. В шестом разделе приведены общие сведения об эндокринной системе, затрагиваются вопросы взаимодействия двух регуляторных систем организма – нервной и гуморальной.
Введение
1.1. Содержание предмета «Анатомия ЦНС»
1.2. Методы
1.3. Задачи
1.4. Строение нервной системы
1.5. Анатомическая номенклатура
1.1. Содержание предмета «Анатомия ЦНС»
Классическая анатомия центральной нервной системы представляет собой раздел фундаментальной академической дисциплины - анатомии человека. Анатомия человека рассматривает форму и строение человеческого тела, всех его частей и органов в связи с их функцией, развитием и влиянием окружающей организм среды. Анатомия центральной нервной системы дает представление об устройстве и топографо-анатомических отношениях мозговых структур с учетом их микроскопического строения, с привлечением современных сведений о функциональном назначении каждой структуры.
Особенностью данного предмета является невозможность формирования правильных представлений об устройстве нервной системы без учета с одной стороны определенных взаимоотношений между организмом и окружающей его средой, с другой стороны - определенных взаимоотношений внутри самого организма. Сам организм человека представляет собой единое целое, сложный комплекс частей и органов, не просто механически соединенных, а находящихся в функциональной взаимосвязи, где всякое изменение в его любой части неизбежно отражается на деятельности всех остальных частей. Поэтому при изучении строения нервной системы необходимо учитывать ее двусторонние связи с другими анатомическими структурами. Важно и влияние химических факторов, которое лежит в основе взаимосвязи двух интегрирующих систем организма – нервной и гуморальной. Эти взаимоотношения достаточно сложны, даже если бы мы рассматривали организм вне связи с окружающей средой. Для человека отношения с окружающей средой еще более усложняются разнообразными социальными взаимоотношениями.
Необходимо помнить, что анатомические особенности нервной системы также определяются и особенностями развития - как индивидуального (онтогенез), регулируемого в основном эпигенетическими факторами – внешними и внутренними, так и эволюционного (филогенез), представляющего собой результат генетических преобразований.
В результате всего вышеперечисленного в рамках курса «Анатомия ЦНС» нам предстоит ответить на следующие вопросы:
как устроена нервная система на микро- и макроскопическом уровнях
каким образом морфологические особенности нервной ткани определяют функции нервных структур
почему именно так организована нервная система человека
Методы
Название «анатомия» происходит от греческого слова anatemno - рассекаю. Это связано с тем, что первым методом анатомического исследования являлось рассечение трупа на части, или препарирование. Хороший анатомический препарат позволяет правильно определить и грамотно описать топографо-анатомические взаимоотношения отделов ЦНС на макроскопическом уровне.
Развитие клеточной биологии, техники микроскопического препарирования: световой, электронной, конфокальной микроскопии - дополнило данные макроанатомии сведениями о клеточном строении нервной системы, об ультраструктурных особенностях нейронов, глиальных клеток; о строении специализированных контактов нейронов – синапсов. Наука, родившаяся на стыке нейроанатомии, нейрофизиологии и нейрохомии, и расширившая наши познания в области организации субклеточных структур нервной ткани, их биофизических свойств, получила название нейробиологии.
Следует сказать несколько слов о методах, не являющихся непосредственно методами анатомии ЦНС, но, безусловно обогативших наши представления о ней. Современное развитие науки и техники позволяет изучать не только иссеченные (посмертные) препараты нервной ткани, появились методы дающие возможность прижизненного изучения мозговых структур. Сюда следует отнести основные методы медицинской диагностики: энцефалографию, эхоэнцефалоскопию, компьютерную томографию и др., дающие представление об изменении структуры и функций нервной ткани при изменении физической нагрузки и развитии заболеваний.
Задачи
Все вышеперечисленные методы позволяют решить следующие задачи:
изучить детали внешнего и внутреннего строения мозговых структур
установить уровни организации и интеграции отдельных нервных элементов и отделов ЦНС в организме
выявить роль различных структур нервной системы в жизнедеятельности животных и человека
Иными словами, все задачи и методы «Анатомии ЦНС» позволяют нам ответить на вопросы «как устроена нервная система организма, почему она устроена именно таким образом?». Ответ на вопрос, «каким образом это работает?» студенты получат, изучив следующий курс – «Физиологию ЦНС».
1.4. Строение нервной системы
Нервная система в соответствии с общей топографией организма разделена на два отдела: центральный и периферический. Центральный отдел составляют те части нервной системы, которые находятся внутри костных образований. Головной мозг – часть ЦНС, заключенная внутри черепа, спинной мозг – часть ЦНС, заключенная внутри позвоночника. К периферическому отделу относят те части нервной системы, которые лежат вне черепа или позвоночника – нервы и нервные узлы.
В соответствии с другой классификацией, в основе которой лежат функциональные особенности нервной регуляции, всю нервную систему (и центральную и периферическую) делят на два других отдела: соматический и вегетативный. Основные отличия этих отделов представим виде схемы:
Нужно отчетливо представлять, что разделение нервной системы на соматическую и вегетативную, равно как и на центральную и периферическую носит условный характер, так как все отделы нервной системы анатомически и функционально связаны друг с другом, являются одним целым (рис. 1). Более того, между центрами головного и спинного мозга и регулируемыми ими органами существует не только прямая, но и обратная связь. Иными словами, из органов, деятельность которых изменяется под влиянием нервных импульсов, посылаемых мозговыми центрами, обратно в мозг поступает информация о характере этих изменений.
Нервная система образована специфической возбудимой тканью – нервной тканью. Основными клетками нервной ткани являются нервные и глиальные клетки.
Нервная клетка (нейрон) специализирована таким образом, что она способна приходить в состояние возбуждения:
принимать определенные формы сигналов
отвечать специальными сигналами
создавать специфические контакты с другими клетками.
Из-за этих свойств нейрон считается структурно-функциональной единицей нервной ткани.
Глиальные (греч. glia - клей) клетки выполняют многие функции в нервной системе, но никогда не проводят нервных импульсов. Различают несколько видов глиальных клеток, для них характерны опорная, трофическая, секреторная, защитная и некоторые другие функции.
Центральная нервная система - головной и спинной мозг – состоят из огромного количества нервных клеток, их отростков, а также из нейроглии, образующих серое и белое мозговое вещество. Серое вещество представляет собой скопление нервных клеток с отходящими от них отростками. Белое вещество – скопление нервных волокон. Нейроглия входит в состав как серого, так и белого вещества. В головном мозге, в разных его отделах серое вещество расположено неодинаково. В больших полушариях и в мозжечке серое вещество образует снаружи сплошной слой, называемый корой (cortex). Под корой находится белое вещество, а в нем – отдельные скопления серого вещества – ядра (nucleus). В других отделах головного мозга белое вещество расположено снаружи, а серое вещество в виде ядер внутри. Некоторые отделы состоят преимущественно из ядер. В спинном мозге белое вещество всегда снаружи, а серое внутри, также имеет ядерное строение. Ядра серого вещества играют роль различных центров головного и спинного мозга, регулируют функции органов (центр слюноотделения, центр дыхания, центр глотания и т.д.). Пучки нервных волокон белого вещества связывают одни отделы мозга с другими и выполняют проводниковую функцию.
Периферическая нервная система представлена черепными и спинномозговыми нервами и их окончаниями. Нервы состоят из пучков нервных волокон, окруженных общими оболочками. Многие нервы и их разветвления имеют помимо нервных волокон и нервные узлы – ганглии (ganglion). Они состоят из нейронов, отростки которых входят в состав данных нервов. Нервы дают ветви ко всем органам нашего тела, или как принято говорить иннервируют их. Нервные окончания могут быть образованы с чувствительными клетками, они получают название рецепторных (или афферентных), и с двигательными – эффекторные (или эфферентные). Сочетание рецепторных, нервных и эффекторных компонентов формируют разные по уровню сложности организации рефлекторные дуги.
Анатомическая номенклатура
Для обозначения взаимного расположения структур внутри организма анатомы пользуются специальными терминами, основанными на латинских названиях (рис. 2; приложение 1):
плоскость, проходящая вдоль по середине тела, делящая его на правую и левую половины, называется сагиттальной (sagittalis)
плоскость, идущая также идущая вдоль по середине тела, но перпендикулярно сагиттальной, называется фронтальной (она параллельна лбу – frontis)
структуры, расположенные на спинной стороне тела называются дорсальными (dorsalis – спинной)
структуры, расположенные на брюшной стороне – вентральными (ventralis – брюшной)
структуры, лежащие по центру тела, близко к сагиттальной плоскости – медиальными (medialis – срединный)
структуры, лежащие сбоку от сагиттальной плоскости – латеральными (lateralis – боковой)
самые верхние точки структур называются апикальными (apicalis – верхушечный)
структуры, лежащие в основании – базальными (basalis)
направление к верхней части тела, к головной его части – ростральное (rostrum – клюв)
направление к нижней части тела – каудальное (caudalis – хвостовой)
По отношению к протяженным органам – конечностям, нервам и т.п. используются следующие термины:
проксимальный – близкий от начала отхождения (от туловища, от спинного или головного мозга)
дистальный - отдаленный
Раздел 1
НЕРВНАЯ ТКАНЬ
Нейроны – специализированные возбудимые клетки нервной системы
2.1. Структура нейрона
2.2. Ультраструктурные особенности нервных клеток
2.2.1. Клеточная мембрана
2.2.2. Цитоплазма и клеточные органеллы
2.3. Классификация нейронов
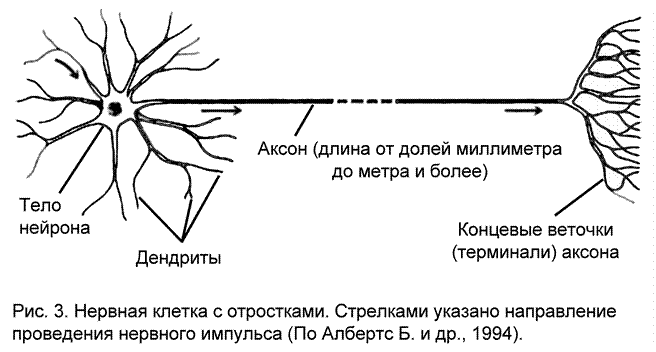 2.1. Структура нейрона
2.1. Структура нейрона
Нейрон – это нервная клетка со всеми ее отростками. Специфическая функция нервных клеток состоит в том, что они приходят в состояние возбуждения под влиянием раздражения и проводят раздражение в виде нервных импульсов. В соответствии с разными научными исследованиями в ЦНС человека насчитывается до 1011 разных нейронов. Все нейроны имеют общие морфологические признаки: они состоят из тела и специализированных отростков – аксона (или по-другому – нейрита) и дендритов (рис. 3).
Следует сразу отметить, что такая морфологическая поляризация (дендрит – сома – аксон) связана с их функциональной поляризацией. Это проявляется в том, что только аксон клетки имеет на своих концевых разветвлениях структуры, предназначенные для передачи активности на другие клетки. Аксоны каждого нейрона, подходя к другим клеткам, ветвятся, образуя многочисленные окончания на дендритах этих клеток, их телах и на конечных разветвлениях – терминалях аксонов. На теле крупной нервной клетки, могут располагаться тысячи нервных окончаний, образованных отростками других нейронов (рис. 4 а). А одно нервное волокно, в свою очередь, может образовывать множество таких контактов на других клетках (рис. 4 б). Такие контакты между клетками называются синаптическими, или синапсами (от греч. sinapsis - соединение).
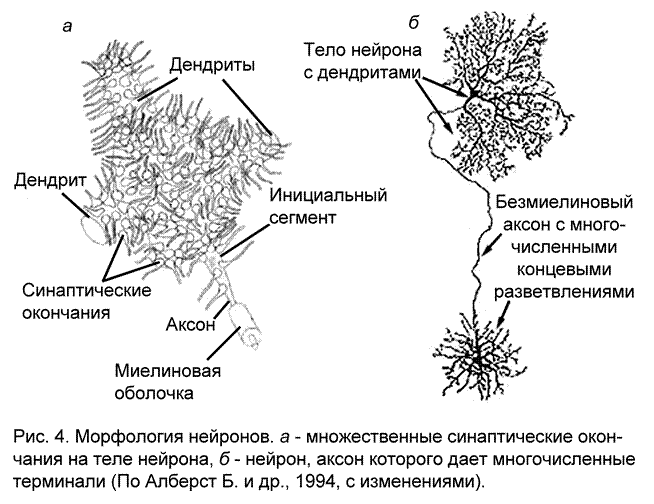 Тело нейрона содержит все компоненты клеточных структур. Оно выполняет по отношению к своим отросткам, прежде всего трофическую функцию, способно генерировать нервные импульсы. Основное значение в нейроне имеет участок, где возникает нервный импульс – инициальный сегмент (рис. 4 а). Именно здесь зарегистрирована самая высокая активность (уровень возбудимости) нейронов.
Тело нейрона содержит все компоненты клеточных структур. Оно выполняет по отношению к своим отросткам, прежде всего трофическую функцию, способно генерировать нервные импульсы. Основное значение в нейроне имеет участок, где возникает нервный импульс – инициальный сегмент (рис. 4 а). Именно здесь зарегистрирована самая высокая активность (уровень возбудимости) нейронов.
Дендриты (от греч. dendron – дерево, т.е. «древовидные отростки») – это короткие, сильноветвящиеся отростки, являющиеся чувствительной (рецепторной) областью нейрона. Все вместе они составляют конвергентную (от лат. convergentio - сходиться) систему сбора информации, которая к ним поступает через синапсы от других нейронов или от специализированных чувствительных клеток - рецепторов.
Аксон (от греч. acsis – отросток, по-другому он называется нейрит) – это длинный отросток, несущий импульсы от тела нервной клетки, является двигательной (эфферентной) областью нейрона. В отличие от дендритов, которых у клетки может быть множество, аксон всегда один. С морфологической точки зрения аксоны – длинные, гладкие и разветвляются менее чем дендриты. Ответвления аксона называют коллатералями.
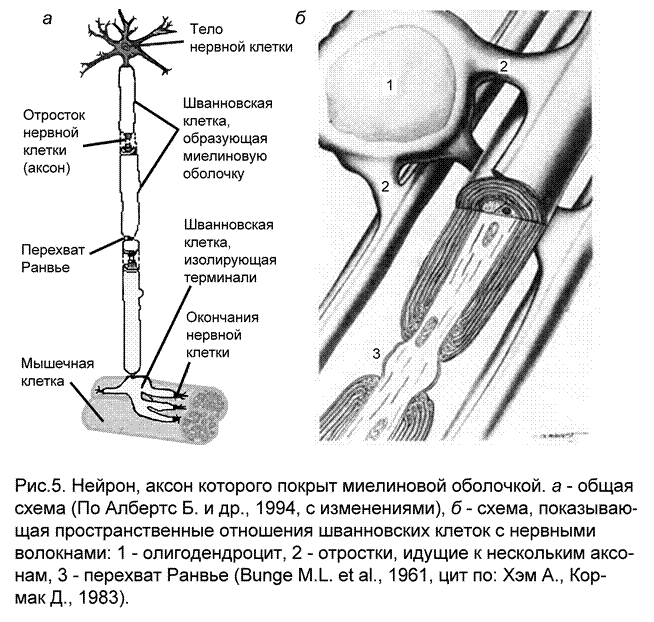 Как пишет чешский нейроанатом Станислав Немечек: « …Более точным является обозначение «нейрит», так как оно лучше выражает противоположность к обозначению «дендрит». Наименование «осевой цилиндр» (сокращенно «аксон») логически следовало бы употреблять только тогда, когда нейрит не обнажен, а находится внутри специальной оболочки, являясь ее осью. Аксон вместе с глиальной оболочкой можно назвать нервным волокном. Англо-американское обозначение обнаженных нейритов как «аксоны», не точно. Однако оно настолько прижилось, что в настоящее время уже трудно обойтись без него. Так что мы вынуждены его тоже применять…».
Как пишет чешский нейроанатом Станислав Немечек: « …Более точным является обозначение «нейрит», так как оно лучше выражает противоположность к обозначению «дендрит». Наименование «осевой цилиндр» (сокращенно «аксон») логически следовало бы употреблять только тогда, когда нейрит не обнажен, а находится внутри специальной оболочки, являясь ее осью. Аксон вместе с глиальной оболочкой можно назвать нервным волокном. Англо-американское обозначение обнаженных нейритов как «аксоны», не точно. Однако оно настолько прижилось, что в настоящее время уже трудно обойтись без него. Так что мы вынуждены его тоже применять…».
Аксон, покрытый глиальной оболочкой, принято называть нервным волокном. Выделяют два вида нервных волокон:
безмиелиновые нервные волокна (рис. 4 б) покрыты одним слоем глиальных клеток, между которыми существуют щелевидные пространства. Клеточная мембрана аксона при таком способе покрытия глиальными компонентами по всей длине имеет контакт с окружающей межклеточной средой. Безмиелиновые нервные волокна способны генерировать нервные импульсы на всем своем протяжении.
миелиновые нервные волокна (рис. 4 а, 5) покрыты слоями глиальных клеток, которые формируют миелиновую оболочку. Миелиновая оболочка состоит из белково-липидного комплекса – миелина и образуется вследствие многократного обертывания аксона клетками глии. В центральной нервной системе эту роль выполняют олигодендроциты, в периферической – шванновские клетки. По ходу миелиновой оболочки встречаются участки без миелина, они называются перехватами Ранвье и соответствуют границам между миелинизироваными участками. В этих участках внеклеточная жидкость непосредственно контактирует с мембраной аксона. 
|
Волокна, покрытые миелиновой оболочкой, называются мякотными, а волокна, не покрытые миелином, получили название безмякотных. Мякотные нервные волокна входят в состав чувствительных и двигательных нервов, снабжающих органы чувств и скелетную мускулатуру, они также встречаются и в вегетативной нервной системе. Безмякотные волокна принадлежат в основном к симпатической нервной системе.
Выделяют следующие группы волокон:
группа А ( α, b, g, d) – имеют самую толстую миелиновую оболочку и самую высокую скорость проведения нервного импульса
группа В – миелиновая оболочка менее выражена
группа С – без миелиновой оболочки, скорость проведения нервного импульса низка, непрямую зависит от диаметра нервного волокна.
Дата: 2018-11-18, просмотров: 478.