Молекулы ДНК являются полимерами, мономерами которых являются дезоксирибонуклеотиды, образованные:
© остатком пятиуглеродного сахара — дезоксирибозы;
© остатком одного из азотистых оснований:
¨ пуриновых — аденина, гуанина;
¨ пиримидиновых — тимина, цитозина;
© остатком фосфорной кислоты.
|
Структура молекулы ДНК |
ДНК представляет собой двойную спираль. Ее молекула образована двумя полинуклеотидными цепями, спирально закрученными друг около друга, и вместе вокруг воображаемой оси (рис. 273). Цепи ДНК антипараллельны (разнонаправлены), то есть против 3'-конца одной цепи находится 5'-конец другой. На периферию молекулы обращен сахаро-фосфатный остов, образованный чередующимися остатками дезоксирибозы и фосфатными группами. Внутрь молекулы обращены азотистые основания.
Диаметр двойной спирали ДНК — 2 нм, шаг общей спирали, на который приходится 10 пар нуклеотидов — 3,4 нм. Длина молекулы — до нескольких десятков и даже сотен микрометров. Молекулярный вес составляет десятки и сотни миллионов (для двойной спирали). В ядре клетки человека общая длина ДНК около 2м.

Трехмерная модель пространственного строения молекулы ДНК в виде двойной спирали была предложена в 1953 г. американским биологом Дж.Уотсоном и английским физиком Ф.Криком (рис. 274). За свои исследования они были удостоены Нобелевской премии.
Полинуклеотидные цепи в молекуле ДНК удерживаются друг около друга благодаря возникновению водородных связей между азотистыми основаниями. Спаривание нуклеотидов не случайно, в его основе лежит принцип комплементарного взаимодействия пар оснований: против аденина одной цепи всегда располагается тимин на другой цепи, а против гуанина одной цепи — всегда цитозин другой, то есть аденин комплементарен тимину, а гуанин — цитозину (рис. 275). Комплементарностью называют способность нуклеотидов к избирательному соединению друг с другом.
|
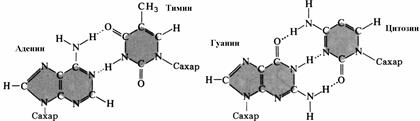 Рис. 275. Спаривание азотистых оснований.
Рис. 275. Спаривание азотистых оснований.
Комплементарность обеспечивается:
© взаимодополнением пространственных конфигураций молекул азотистых оснований;
© количеством водородных связей, возникающих между азотистыми основаниями (три водородные связи между гуанином и цитозином и две водородные связи между аденином и тимином).
Комплементарность пуриновых и пиримидиновых азотистых оснований обеспечивает одинаковое по всей длине двойной спирали расстояние между цепями.
Э.Чаргафф, обследовав огромное количество образцов тканей и органов различных организмов, выявил следующую закономерность: в любом фрагменте ДНК содержание остатков гуанина всегда точно соответствует содержанию цитозина, а аденина — тимину. Это положение получило название "правила Чаргаффа":
А + Г
А = Т; Г = Ц или ——— = 1
Ц + Т
Дж.Уотсон и Ф.Крик воспользовались этим правилом при построении модели молекулы ДНК.
Последовательность нуклеотидов одной цепи определяет последовательность нуклеотидов другой, поэтому две цепи молекулы ДНК комплементарны друг другу.
|
Самоудвоение молекулы ДНК |
Одним из уникальных свойств молекулы ДНК является ее способность к самоудвоению — воспроизведению точных копий исходной молекулы. Благодаря этой способности молекулы ДНК, осуществляется передача наследственной информации от материнской клетки дочерним во время деления. Процесс самоудвоения молекулы ДНК называют репликацией.
Репликация — сложный процесс, идущий с участием ферментов (ДНК-полимераз) (рис. 276). Репликация осуществляется полуконсервативным способом, то есть под действием ферментов молекула ДНК раскручивается и около каждой цепи, выступающей в роли матрицы, по принципу комплементарности достраивается новая цепь. Таким образом, в каждой дочерней ДНК одна цепь является материнской, а вторая — вновь синтезированной. Раскручивание молекулы происходит на небольшом отрезке (несколько десятков нуклеотидов), называемом репликативной вилкой. После окончания синтеза дочерних цепей ДНК на данном участке и соединения их с материнскими раскручивается новый отрезок, и цикл репликации повторяется. Таким образом, репликативная вилка перемещается вдоль молекулы, пока не дойдет до точки окончания синтеза.
|
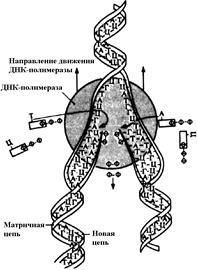 Рис. 276. Репликация ДНК.
Рис. 276. Репликация ДНК.
Во время репликации энергия молекул АТФ не расходуется, так как для синтеза дочерних цепей при репликации используются не дезоксирибонуклеотиды (содержат один остаток фосфорной кислоты), а дезоксирибонуклеозидтрифосфаты (содержат три остатка фосфорной кислоты). При включении дезоксирибонуклеозидтрифосфатов в полинуклеотидную цепь два концевых остатка отщепляются, и освободившаяся энергия используется на образование сложноэфирной связи между нуклеотидами.
Рибонуклеиновые кислоты
Молекулы РНК являются полимерами, мономерами которых являются рибонуклеотиды, образованные:
¨ остатком пятиуглеродного сахара — рибозы;
¨ остатком одного из азотистых оснований:
¨ пуриновых — аденина, гуанина;
¨ пиримидиновых — урацил, цитозина;
¨ остатком фосфорной кислоты.
Структурная организация РНК
Молекула РНК представляет собой неразветвленный полинуклеотид, имеющий третичную структуру. В отличие от ДНК, она образована не двумя, а одной полинуклеотидной цепочкой. Однако ее нуклеотиды также способны образовывать водородные связи между собой, но это внутри–, а не межцепочечные соединения комплементарных нуклеотидов. Цепи РНК значительно короче цепей ДНК.
Информация о структуре молекулы РНК заложена в молекулах ДНК. Синтез молекул РНК происходит на матрице ДНК с участием ферментов РНК-полимераз и называется транскрипцией. Последовательность нуклеотидов в РНК комплементарна кодирующей цепи ДНК и идентична, за исключением замены тимина на урацил, некодирующей цепи.
Если содержание ДНК в клетке относительно постоянно, то содержание РНК сильно колеблется. Наибольшее количество РНК в клетках наблюдается во время синтеза белка.
Существует три основных класса рибонуклеиновых кислот:
© информационная (матричная) РНК — иРНК;
© транспортная РНК — тРНК;
© рибосомальная РНК — рРНК.
|
Информационная РНК |
Наиболее разнообразный по размерам и стабильности класс. Все они являются переносчиками генетической информации из ядра в цитоплазму. Они служат матрицей для синтеза молекулы белка, т.к. определяют аминокислотную последовательность первичной структуры белковой молекулы.
На долю иРНК приходится до 5% от общего содержания РНК в клетке.
|
Транспортная РНК |
Молекулы транспортных РНК содержат обычно 75-86 нуклеотидов. Молекулярная масса молекул тРНК » 25000. Молекулы тРНК играют роль посредников в биосинтезе белка — они доставляют аминокислоты к месту синтеза белка, в рибосомы. В
|
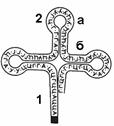 Рис. 277. Строение тРНК:
1 — акцепторное плечо; 2 — антикодоновое плечо (а — петля; б — "стебель").
Рис. 277. Строение тРНК:
1 — акцепторное плечо; 2 — антикодоновое плечо (а — петля; б — "стебель").
Каждый вид тРНК имеет характерную только для него последовательность нуклеотидов. Однако у всех молекул имеется несколько внутримолекулярных комплементарных участков, благодаря наличию которых все тРНК имеют третичную структуру, напоминающую по форме лист клевера (рис. 277).
Молекулы всех тРНК имеют четыре основных плеча:
© акцепторное;
© антикодоновое;
© два боковых.
Каждое плечо состоит из "стебля", образованного комплементарными парами оснований, и петель из неспаренных оснований.
Акцепторное плечо через 3¢-гидроксильную группу аденозильного остатка связывает тРНК с аминокислотой. Антикодоновое плечо содержит триплет нуклеотидов (антикодон), комплементарный кодону иРНК.
|
Рибосомная РНК |
На долю рибосомальной РНК (рРНК) приходится 80-85% от общего содержания РНК в клетке. Рибосомная РНК состоит из 3-5 тыс. нуклеотидов. В комплексе с рибосомными белками рРНК образует рибосомы — органеллы, на которых происходит синтез белка.
Основное значение рРНК состоит в том, что она обеспечивает первоначальное связывание иРНК и рибосомы и формирует активный центр рибосомы, в котором происходит образование пептидных связей между аминокислотами в процессе синтеза полипептидной цепи.
Дата: 2018-11-18, просмотров: 422.