Пигментные наборы фотосинтезирующих организмов позволяют им использовать весь диапазон длин волн падающей на Землю солнечной энергии. Обращает внимание большое различие в спектрах поглощения у представителей разных групп фотосинтезирующих организмов и прежде всего существенные сдвиги в максимумах поглощения хлорофиллов в красной области спектра. Несомненно экологическое значение этого явления, позволяющего избегать конкуренции за свет между разными группами фотосинтезирующих организмов. Что же касается эволюции спектров поглощения хлорофиллов, то очевидна тенденция к перемещению в более коротковолновую часть спектра с более высоким энергетическим уровнем.
Световая фаза фотосинтеза
5.1 Фотофизический этап. Электронно-возбужденное состояние пигментов. Представление о фотосинтетической единице. Антенные комплексы. Реакционные центры. Преобразование энергии в реакционном центре
На световой фазе фотосинтеза происходит поглощение света молекулами хлорофилла а с участием дополнительных пигментов (хлорофилла b, каротиноидов, фикобилинов) и трансформация энергии света в химическую энергию АТФ и восстановленного НАДФ. Все эти процессы осуществляются в фотохимически активных мембранах хлоропластов и представляют собой сложную систему фотофизических, фотохимических и химических реакций. В состав ламелл хлоропластов входят пять многокомпонентных белковых комплексов: светособирающий комплекс, фотосистемы I и II, цитохромный комплекс, включающий цитохромы b6 и f5 и АТФазный комплекс, участвующий в синтезе АТФ. Все они функционируют, взаимодействуя друг с другом.
Две фотосистемы. Впервые идею о существовании в хлоропластах двух фотосистем высказал Р. Эмерсон (1957), изучая влияние света на квантовый выход фотосинтеза у хлореллы. Под квантовым выходом фотосинтеза понимается количество выделившегося 02 или связанного С02 на 1 квант поглощенной энергии. Было показано, что квантовый выход высок при освещении хлореллы красными лучами с длиной волны 660-680 нм. Использование красного света с большей длиной волны приводило к снижению квантового выхода, а при 700 нм фотосинтез почти прекращался, хотя эта часть спектра еще поглощается хлорофиллом. Однако если хлореллу одновременно освещали коротковолновым (650 нм) и длинноволновым (700 нм) красным светом, то суммарный эффект (Y) был выше, чем при действии каждого красного света в отдельности.
Это явление получило название эффекта усиления Эмерсона. Отсюда возникло предположение, что в хлоропластах взаимодействуют две пигментные системы.
Предположение Эмерсона о двух пигментных системах подтвердилось в последующих работах, в которых из мембран хлоропластов с помощью детергентов (поверхностно-активных веществ, диссоциирующих гидрофобные связи) дифференциального центрифугирования в градиенте плотности сахарозы и других приемов удалось выделить и изучить белковые комплексы фотосистемы I (ФС I) и фотосистемы II (ФС II).
В состав ФС I в качестве реакционного центра входит димер пигмента П700 (хлорофилл а с максимумом поглощения 700 нм), а также хлорофиллы а675-695, играющие роль антенного компонента фотосистемы I. Первичным акцептором электронов в этой системе является мономерная форма хлорофилла а695 (Ах), вторичными акцепторами — А2 и Ав (железосерные белки — FeS). Комплекс ФС I под действием света восстанавливает водорастворимый FeS-белок ферредоксин (Фд) и окисляет медьсодержащий водорастворимый белок пластоцианин (Пц).
Белковый комплекс ФС II включает в себя реакционный центр содержащий хлорофилл а П680, а также антенные пигменты — хлорофиллы a670-673. Первичным акцептором электронов в этой фотосистеме выступает феофетин а (Фф), передающий электроны на первичный пластохинон, ассоциированный с FeS (QA) и дальше на вторичный пластохинон (Qb). В состав ФС II входят также белковый комплекс S-системы, окисляющий воду, и переносчик электронов Z, связанный с системой S и служащий донором е для П680. Этот комплекс функционирует с участием марганца, хлора и кальция. В ФС II локализован цитохром b559. ФС II восстанавливает пластохинон (PQ) и окисляет воду с выделением 02 и протонов.
Связующим звеном между ФС II и ФС I служат пул (фонд) пластохинонов, белковый цитохромный комплекс b6 — f5 и пластоцианин.
В ходе эволюции ФС I возникла раньше. Она функционирует у ныне существующих фотосинтезирующих бактерий, фотосинтез которых осуществляется без разложения воды и выделения 02 (фоторедукция). В этом случае функцию доноров электронов для световой фазы фотосинтеза выполняют легко окисляемые соединения, такие, как H2S, Н2, СН4 и др.
Антенные комплексы и миграция энергии в пигментных системах. Эмерсон и Арнольд показали, что в хлоропластах растений на каждый реакционный центр приходится от 200 до 400 молекул хлорофилла, функции которых связаны с поглощением света и передачей энергии возбужденного состояния на реакционные центры. Молекулы хлорофилла а и дополнительные пигменты — хлорофилл b, каротиноиды, фикобилины — входят в состав антенных или светособирающих комплексов (ССК). Из ламелл хлоропластов выделены светособирающий белковый комплекс с хлорофиллами а и b (ССКa-Ь ), тесно связанный с ФС II, и антенные комплексы, непосредственно входящие в фотосистемы I и II (фокусирующие антенные компоненты фотосистем).
В ССКa-Ь присутствуют хлорофилл а в двух или трех формах с максимумами поглощения между 660 и 675 нм, хлорофилл b с максимумом поглощения 650 нм и каротиноиды. В каждом ССКa-b содержится от 120 до 240 молекул хлорофиллов, причем отношение хлорофилла а к хлорофиллу b составляет 1,2—1,4. Половина белка тилакоидов и около 60% общего количества хлорофилла локализовано в ССК. У сине-зеленых и красных водорослей, у которых хлорофилл b отсутствует, роль ССК выполняют фикобилисомы, в состав которых входят фикобилины.
Антенный белковый комплекс ФС II содержит 40 молекул хлорофиллов а с максимумами поглощения 670 — 683 нм на один П680 и β-каротин.
Антенный белковый комплекс ФС I состоит из хромопротеинов, содержащих 110 молекул хлорофиллов а с максимумами поглощения 680 — 695 нм на один П700, из них 60 молекул — компоненты антенного комплекса самой фотосистемы, а 50 входят в состав комплекса, который можно рассматривать как ССК ФС I. Антенный комплекс ФС I также содержит β-каротин.
Хромопротеины антенных комплексов не обладают фотохимической и энзиматической активностью. Если бы каждая молекула хлорофилла преобразовывала поглощенную ею энергию кванта света в фотохимическую реакцию, то такая система была бы крайне нерентабельна. Продолжительность синглетного возбужденного состояния исчисляется 10-12 — 10-9 с, и даже на прямом солнечном свету 1 квант света поглощается молекулой хлорофилла не чаще одного раза за 0,1 с. Большую часть времени молекула хлорофилла «простаивает». Поэтому роль пигментов антенных комплексов состоит в том, чтобы собирать и передавать энергию квантов на небольшое количество молекул реакционных центров П680 и П700, которые и осуществляют фотохимические реакции. Аналогично отдельные капли дождя ударяют в крышу и, сливаясь, создают постоянный ток воды в водостоке.
Передача (миграция) энергии по пигментам антенных комплексов происходит по принципу индуктивного резонанса (без флуоресценции и переноса заряда). Природа индуктивного резонанса сострил в следующем. Каждая молекула хлорофилла, поглотившая квант света и перешедшая в синглетное возбужденное состояние, является молекулярным осциллятором. Возникающее вокруг возбужденной молекулы переменное электрическое поле с определенной частотой колебаний индуцирует осцилляцию диполя (электрон — ядро) соседней молекулы. При этом молекула-донор переходит в основное состояние, а молекула-акцептор — в возбужденное. Условиями для резонансного переноса энергии электронного возбуждения служат малые расстояния между молекулами, не превышающие 10 нм, и перекрытие частот колебаний у двух взаимодействующих молекул.
Флуоресценция каждой молекулы-донора имеет более длинноволновый максимум по сравнению с максимумом ее поглощения (в соответствии с правилом Стокса) и в большей или меньшей степени перекрывает зону поглощения акцепторной молекулы. Миграция энергии осуществляется от коротковолновых пигментов в сторону все более длинноволновых, т. е. пигментов с более низким уровнем синглетного возбужденного состояния.
В антенных комплексах перенос энергии осуществляется в ряду: каротин (400-550 нм)--► хлорофилл b (650 нм)--►хлорофиллы a (660-675 нм)---► П680 (ФС II). Скорость резонансного переноса энергии от молекулы к молекуле 10-10 — 10-9 с, причем эффективность переноса между молекулами хлорофилла достигает 100, а между молекулами каротина и хлорофилла — лишь 40%.
Реакционные центры. Белки, содержащие длинноволновые формы хлорофиллов (П680 в ФС II и П700 в ФС I) в количестве одна молекула на 200 — 400 молекул других хлорофиллов и способные к первичному фотохимическому разделению зарядов, называют реакционными центрами.
Первичное разделение зарядов в реакционных центрах происходит между молекулами хлорофилловой природы и связано с транспортом электронов. Функцию первичного донора электронов выполняет молекула хлорофилла, находящаяся в синглетном возбужденном состоянии. В реакционном центре ФС II, перешнуровывающем мембрану, первичным донором электронов служит П680, а первичным акцептором — феофетин. В реакционном центре ФС I первичный донор электронов— П700, а акцептор — мономерная форма хлорофилла а695 (А1).
Порядок реакций в реакционном центре ФС II определяется продолжительностью того или иного состояния молекул. Молекулы находятся в синглетном возбужденном состоянии 10-10 — 10-9 с. Первичные акцепторы (Фф или А1) получают электроны от своих доноров (П680 или П700 соответственно) за время около 10-12 с. Время обратных рекомбинаций от Фф- на П680+ и от A1- на П700+ во много раз больше (10-6 с). То же самое относится к последующим парам взаимодействующих молекул. Поэтому весь ход реакций резко сдвинут вправо. В реакционных центрах энергия возбужденного состояния хлорофилла преобразуется в энергию разделенных зарядов, т. е. превращается в химическую энергию.
Фотохимический этап. Электронно-транспортная цепь фотосинтеза. Представления о функционировании двух фотосистем. Фотофосфорилирование. Системы фотоокисления воды и выделения кислорода при фотосинтезе. Связь фотосинтетической ассимиляции С02 с фотохимическими реакциями
Нециклический и циклический транспорт электронов.
Для восстановления одной молекулы NADP+ в процессе фотосинтеза необходимы два электрона и два протона, причем донором электронов является вода. Фотоиндуцированное окисление воды осуществляет ФС II, восстановление NADP+ — фотосистема I. Следовательно, эти две фотосистемы должны функционировать во взаимодействии.
Исходя из данных о квантовом выходе фотосинтеза (необходимы 8 квантов света для выделения одной молекулы 02) из состава компонентов, входящих в ФС I и ФС II, и величин их окислительно-восстановительного потенциала была разработана схема последовательности реакций в световой фазе фотосинтеза.
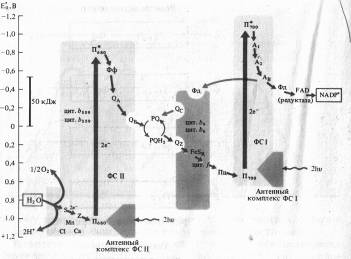
Из-за сходства с буквой Z эта схема, получила название Z-схемы. Впервые принцип Z-схемы был предложен Р. Хиллом и Ф. Бендаллом (1960) и экспериментально подтвержден работами Л. Дюйзенса (1961). В настоящее время Z-схема, или схема нециклического транспорта электронов при фотосинтезе является общепризнанной и непрерывно пополняется новыми деталями.
В ФС II димер П680, поглотив энергию, эквивалентную 2 квантам коротковолнового красного света, и перейдя в синглетное возбужденное состояние, отдает 2 электрона феофетину (Фф). От Фф электроны, теряя энергию, последовательно передаются на пластохиноны ФС 11 - QA и QB, на пул липидорастворимых молекул пластохинона (PQ), переносящих через липидную фазу мембраны электроны и протоны, на железосерный белок FeSK и цитохром цитохромного комплекса b6 — f, восстанавливая Cu-содержащий белок пластоцианин (Пц).
Вакантные места («дырки») в Пб80 заполняются двумя электронами из содержащего Мп переносчика электронов Z, который в свою очередь восстанавливается с участием системы S. Окисленный белковый комплекс S связывает воду и восстанавливается за счет электронов воды. Для осуществления этой реакции в белковом комплексе S необходимы Мп и С1-, а также Са2 + .
Однако описанная цепь реакций резко замедляется, если не возбуждена ФС I, так как в этом случае весь Пц переходит в восстановленное состояние. При возбуждении П700 в реакционном центре ФС I энергией, эквивалентной 2 квантам длинноволнового красного света, 2 электрона захватываются мономерной формой хлорофилла a (A1) и затем последовательно передаются переносчикам электронов А2 и Ав (железосерные белки FeS), ферредоксину (водорастворимый FeS-белок) и ферредоксин: NADP-оксидоредуктазе с FAD в качестве кофактора. Наконец, редуктаза восстанавливает NADP + .
На вакантные места в П700+ переходят электроны с Пц, и нециклическая цепь переноса электронов таким образом замыкается. Z-схема убедительно объясняет «эффект усиления Эмерсона», так как только при совместном использовании коротковолновых (ФС II) и длинноволновых (ФС I) лучей красного света интенсивность фотосинтеза будет оптимальной.
Энергия, освобождающаяся при движении электронов от П680 до П700, используется для синтеза АТФ из AДФ и неорганического фосфата (фотофосфорилирование).
Наряду с нециклическим в мембранах хлоропластов функционирует циклический транспорт электронов, включающий в себя только ФС I и комплекс цитохромов Ь6 — f. В этом случае возбужденные молекулы П700 последовательно отдают электроны на А1, А2, АB, Фд, PQ, цитохром b6, FeSR, цитохром f, Пц и, наконец, П700. При этом НAДФ+ не восстанавливается. Освобождающаяся энергия используется для фосфорилирования AДP.
Интересно отметить, что комплекс цитохромов Ь6 — f функционирует в системе транспорта электронов при фотосинтезе аналогично комплексу III (цит. Ь, C1) в митохондриях.
Разница в уровнях энергии между П680 и П700 (> 50 кДж) вполне достаточна для фосфорилирования AДФ, так как величина высокоэнергетической фосфатной связи АТФ равна 30,6 кДж/моль (7,3 ккал). Перепады энергии в ФС I еще значительнее. Однако само по себе освобождение энергии при нециклическом и циклическом транспорте электронов не объясняет, каким образом осуществляется фотофосфорилирование.
Механизм фосфорилирования AДФ, сопряженного с деятельностью ЭТЦ , объясняет хемиосмотическая теория, разработанная английским биохимиком П. Митчеллом (1961 — 1966). Для объяснения процессов фотофосфорилирования эта теория была впервые использована А. Ягендорфом (1967).
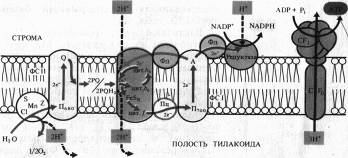
Сущность хемиосмотической теории состоит в следующем. Цепь переносчиков электронов и прогонов, действующая в соответствии с окислительно-восстановительным градиентом, перешнуровывает мембрану таким образом, что трансмембранный перенос е- и Н+ в одну сторону чередуется с переносом в обратную сторону только е-. В результате функционирования такого механизма (Н + -помпы) по одну сторону мембраны накапливается избыток Н+ и возникает электрохимический (т. е. электрический и концентрационный) потенциал ионов Н+, который служит формой запасания энергии. Обратный пассивный ток ионов Н+ через протонный канал Н+-АТФазы, получивший название сопрягающего фактор CF1, сопровождается образованием высокоэнергетической фосфатной связи АТФ.
На рис. 6.1 представлена упрощенная схема расположения переносчиков в мембране тилакоидов, объясняющая механизм фотофосфорилирования. На внутренней стороне мембраны тилакоида под действием света возбуждаются П680 и П700. Электроны от П680 захватываются акцепторами е- на наружной стороне и передаются на окисленную форму пластохинона (PQ) — липидорастворимого переносчика прогонов и электронов, выполняющего функцию челнока. Принимая два электрона, 2PQ диффундируют к цитохромному комплексу и захватывают из стромы 2Н + . Электроны с 2PQH2 поступают в цепь: F.eSR -> цит. f -> Пц,-> П700, а ионы Н + попадают в полость тилакоида. Вторая пара ионов Н+ освобождается там же при фотоокислении воды. В ФС I электроны П700 захватываются акцептором А, передаются ферредоксину и затем НAДФ+ на наружной стороне мембраны, причем 1Н+ используется на восстановление НAДФ.
Таким образом, из среды, окружающей тилакоид, при поглощении пигментами квантов света исчезают, а во внутренней полости тилакоида появляются протоны. В результате на мембране возникает электрохимический потенциал ионов Н + , который затем используется для фосфорилирования AДФ. Этот процесс называется нециклическим фотофосфорилированием.
При циклическом фотофосфорилировании, когда функционирует только ФС I, электроны от Фд поступают на цитохромный комплекс с использованием пула PQ, который действует как переносчик электронов и протонов. Затем электроны через цитохром f и Пц возвращаются на основной энергетический уровень в П700, а протоны поступают в полость тилакоида.
Белковые комплексы, участвующие в фотохимических реакциях фотосинтеза, в мембранах хлоропластов распределены неравномерно. Наибольшие различия наблюдаются между теми мембранами, которые плотно состыкованы друг с другом в гранах и теми участками мембран, которые контактируют со стромой (закругленные участки тилакоидов гран и мембраны тилакоидов стромы).
Светособирающий комплекс и пигмент-белковый комплекс ФС II находятся в основном в мембранах, плотно контактирующих друг с другом, причем ССК играет особую роль. В адгезии тилакоидных мембран.
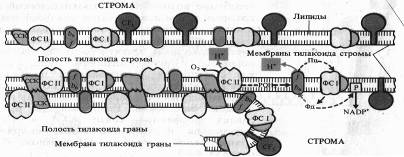
Показано, что у мутанта хламидомонады, лишенного белка, с которым связан хлорофилл Ь, граны не образуются. ФС I со своим светособирающим комплексом преимущественно, а АТФазный комплекс (CF1 + CF0), как правило, находятся на несостыкованных участках мембран. Комплекс цитохромов b6 — f равномерно распределен как в состыкованных, так и в не состыкованных мембранах. Такое распределение белковых комплексов, участвующих в световой фазе фотосинтеза, ставит вопрос о способе их взаимодействия. Показано, что это взаимодействие осуществляется с помощью легкоподвижного липофильного пластохинона PQ в липидной фазе и благодаря перемещению водорастворимого пластоцианина вдоль внутренних поверхностей ламелл и водорастворимого ферредоксина вдоль их наружных поверхностей.
К латеральным перемещениям вдоль мембраны способны и сами белковые комплексы. На это передвижение влияет величина их электрического заряда. Например, восстановление пластохинона PQ фотосистемой II приводит к активации киназы и фосфорилированию ССКа-ь. Фосфорилирование ССК увеличивает его отрицательный заряд, что способствует перемещению ССК в стромальную область мембраны и возрастанию миграции поглощенной энергии света к ФС I. Возросшая фотохимическая активность ФС I усиливает окисление PQ, что, в свою очередь, приводит к инактивации киназы, фосфатаза же дефосфорилирует ССК. Таким образом, обратимое фосфорилирование ССК представляет собой петлю обратной связи в системе взаимной регуляции активности ФС I и ФС II. Механизм такой регуляции включает латеральный транспорт белковых комплексов в мембранах хлоропластов.
6 Метаболизм углерода при фотосинтезе (темновая фаза)
В результате фотохимических реакций в хлоропластах создается необходимый уровень АТР и NADPH. Эти конечные продукты световой фазы фотосинтеза стоят на входе в темновую фазу, где С02 восстанавливается до углевода:
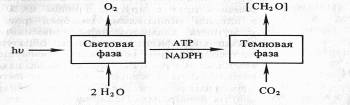
Однако сами по себе АТФ и НAДФH не в состоянии восстановить С02. Очевидно, и темновая фаза фотосинтеза — сложный процесс, включающий большое количество реакций. Кроме того, существуют различные пути восстановления С02. В настоящее время известны так называемые С3-путь и С4-путь фиксации С02, фотосинтез по типу толстянковых (САМ-метаболизм) и фотодыхание. Рассмотрим каждый из этих путей в отдельности.
Дата: 2019-07-30, просмотров: 259.