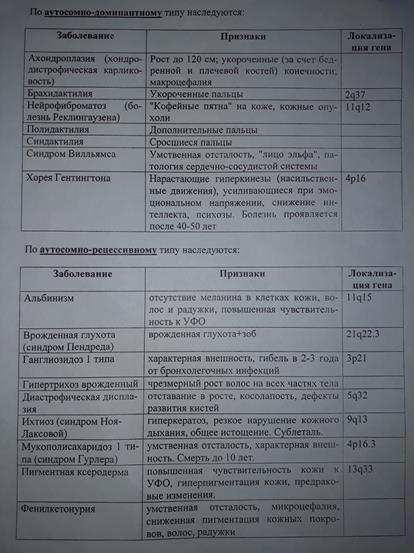
84. Множественный аллелизм (на примере групп крови у человека по системе АВО).
} Во всех рассмотренных до сих пор случаях каждый признак контролируется одним геном, который может быть представлен одним из двух аллелей.
} Известно, однако, немало примеров, когда один признак проявляется в нескольких разных формах, контролируемых тремя и более аллелями, из которых любые два могут находиться в одних и тех же локусах гомологичных хромосом. В таких случаях говорят о множественных аллелях (или множественных аллеломорфах).
} Примером множественного аллелизма является:
- Серия множественных аллелей окраски глаз у дрозофил,
- Окраски шерсти у кроликов,
- Аллели
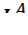 ,
, 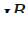 ,
, 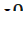 , определяющие группы крови у человека, и т. д.
, определяющие группы крови у человека, и т. д.
} Причиной множественного аллелизма являются случайные мутации, сохраняемые в процессе естественного отбора в генофонде популяции.
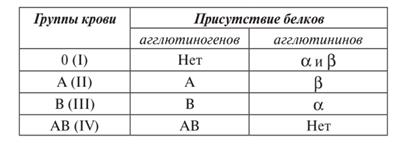
} Группы крови контролируются аутосомным геном. Локус этого гена обозначают буквой I (от слова «изогемагглютиноген»), а три его аллеля – буквами A , B и 0. Аллели A и B доминантны в одинаковой степени, а аллель 0 рецессивен по отношению и к тому, и к другому.
} При гомозиготностиIA IA эритроциты имеют только поверхностный антиген А (группа крови А, или II).
} При гомозиготности IB IB эритроциты несут только поверхностный антиген В (группа В, или III).
} В случае гомозиготности 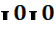 эритроциты лишены А и В антигенов (группа О или I).
эритроциты лишены А и В антигенов (группа О или I).
} В случае гетерозиготности IA 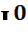 или IB
или IB 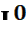 группа крови определяется, соответственно, А (II) или В (III).Эритроциты имеют, соответственно, антигены только А или только В. Это случай полного доминирования.
группа крови определяется, соответственно, А (II) или В (III).Эритроциты имеют, соответственно, антигены только А или только В. Это случай полного доминирования.
При переливании крови, очень важно, чтобы кровь получаемая реципиентом, была совместима с его собственной. В случае несовместимости наблюдается особого рода иммунная реакция. Происходит это потому, что мембрана эритроцитов несет на поверхности гликопротеины – агглютиногены, которые действуют как антигены и реагируют с антителами – агглютининами, содержащимися в плазме крови реципиента.
В результате этого взаимодействия донорские эритроциты агглютинируют, т.е. слипаются друг с другом после связывания с антителами реципиента.
Известно несколько эритроцитарных антигенов, которым соответствуют разные системы групп крови. Наиболее известная из них – система AB0, речь о которой шла ранее.
Лица с группой 0 (I) являются универсальными донорами, поскольку их кровь можно перелить любому человеку. Их эритроциты не несут антигенов, следовательно не могут агглютинировать при контакте с чужой плазмой. Правда в его собственной крови находятся антитела a и b, поэтому при массивных переливаниях опасность агглютинации существует, и группы крови должны совпадать.
Лица с группой AB (IV) – универсальные реципиенты: они могут получать кровь любой группы, так как у них нет агглютиногенов, учитывая сделанную выше оговорку.
85. Цитологические основы законов Менделя.
1. При образовании гамет в результате мейоза в них попадает лишь одна аллель данного гена (это и есть сущность «закона чистоты гамет»).
2. В результате оплодотворения в зиготе оказываются уже две аллели данного гена.
3. В развивающемся из неё новом организме могут иметь место полное или частичное доминирование одной аллели над другой.
Цитологические основы законов Менделя базируются на:
· парности хромосом (парности генов, обусловливающих возможность развития какого-либо признака)
· особенностях мейоза (процессах, происходящих в мейозе, которые обеспечивают независимое расхождение хромосом с находящимися на них генами к разным плюсам клетки, а затем и в разные гаметы)
· особенностях процесса оплодотворения (случайного комбинирования хромосом, несущих по одному гену из каждой аллельной пары)
86. Цитологические основы явления сцепления генов.
Сцепленные гены – это гены, лежащие в одной хромосоме, наследуются вместе. Все гены в одной хромосоме образуют группу сцепления. Число групп сцепления у диплоидных организмов равно гаплоидному набору хромосом. Гомологичные хромосомы образующие на первой стадии мейоза перекресты, способные обмениваться отдельными участками в результате возникающих разрывов и последующих рекомбинаций. Это явление названо кроссинговером (аллели из гомологичных хромосом меняются друг с другом местами). Процесс обмена участками между гомологичными хромосомами приводит к генетической рекомбинации особей, образующихся из гамет с новым сочетанием аллелей называют рекомбинантными. Чем дальше друг от друга на хромосоме расположены гены, тем чаще между ними происходит кроссинговер и тем выше процент появляющихся рекомбинантных особей. На этом явлении основано построение генетических карт – определение последовательности расположения генов в хромосоме и примерного расстояния между ними. Нарушение сцепленного наследования родительских аллелей в результате кроссинговера позволяет говорить о неполном сцеплении в отличии от полного сцепления, наблюдаемого у самцов дрозофилы.
Анализ генетических и цитологических карт позволил сформулировать основные положения хромосомной теории наследственности.
1. Каждый ген имеет определенное постоянное место (локус)
и хромосоме.
2. Гены в хромосомах располагаются в определенной линейной последовательности.
3. Частота кроссинговера между генами прямо пропорциональна расстоянию между ними и обратно пропорциональна силе сцепления.
87. Основные положения хромосомной теории наследственности.
Хромосомная теория наследственности - теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности.
Основные положения
1. Гены находятся в хромосомах; каждая хромосома представляет собой группу сцепления генов; число групп сцепления у каждого вида равно числу пар хромосом.
2. Каждый ген в хромосоме занимает определенное место (локус); гены в хромосомах расположены линейно.
3. Между гомологичными хромосомами происходит обмен аллельными генами.
4. Расстояние между генами (локусами) в хромосоме пропорционально числу кроссинговера между ними.
Дата: 2019-07-31, просмотров: 535.