Переноса
Идентифицировано несколько цитозольных и мембраносвязанных белковых компонентов, необходимых для переноса. Наиболее детально охарактеризованы белковые факторы, участвующие во встраивании белков в эндоплазматический ретикулум млекопитающих (рис.3).
1. Сигнал-распознающая частица (СРЧ). Это растворимый рибонуклеопротеиновый комплекс, состоящий из шести разных белков и молекулы 7S-РНК. СРЧ необходима для инициации переноса. Она связывается с сигнальной последовательностью образующегося полипептида во время его синтеза на рибосоме. Для препролактина, например, константа диссоциации составляет 1нМ. С помощью метода фотохимического сшивания был идентифицирован один из полипептидов (54 кДа), непосредственно взаимодействующий с сигнальной последовательностью предшественника. По некоторым данным, полученным для бесклеточных систем, связывание СРЧ ингибирует трансляцию или вызывает ее задержку. Впрочем, не исключено, что этот феномен является артефактом; во всяком случае, как было показано на модельных опытах, его не обязательно привлекать для объяснения кинетики переноса белков in vivo. Одна из вероятных функции СРЧ состоит в предотвращении неправильного свертывания образующегося полипептида, которое может блокировать перенос (например, из-за экранирования сигнальных последовательностей). Задержка трансляции должна уменьшать вероятность такого ошибочного свертывания и, следовательно, увеличивать эффективность переноса белков.
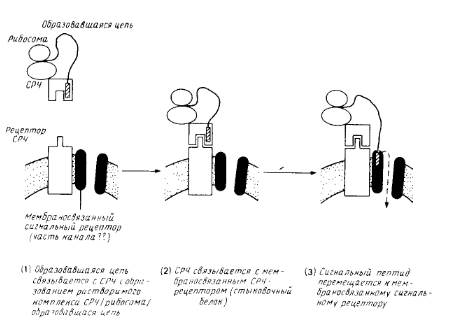
Рис.3. Схематическое изображение ранних стадий котрансляционного переноса полипептида через эндоплазматический ретикулум млекопитающих.
Сигнал-распознающая частица и СРЧ-рецептор (стыковой белок) хорошо охарактеризованы. Мембраносвязывающий сигнальный рецептор изображен как компонент канала через мембрану, но существование таких каналов и роль сигнального рецептора не бесспорны. После образования комплекса между сигнальным пептидом и мембраносвязанным рецептором на стадии 3 СРЧ и стыковочный белок могут диссоциировать и принимать участие в новом цикле, оставляя мембраносвязанную рибосому и образовавшуюся цепь присоединенными к аппарату переноса.
Некоторые небольшие белки (<8,5 кДа) транспортируются в эндоплазматический ретикулум независимо от СРЧ. В их число входят препропептид GLa лягушки, препромеллитин (оба они являются предшественниками секретируемых белков) и пробелок оболочки фага М13. Во всех этих примерах конформация предшественника такова, что белки должны оставаться способными к переносу даже в отсутствии СРЧ и рибосом.
2. Рецептор СРЧ, или стыковочный белок. Комплекс СРЧ/рибосома/образующаяся полипептидная цепь транспортируется в шероховатый эндоплазматический ретикулум, преодолевая энергию сильного взаимодействия между СРЧ, называемым также стыковочным белком. Рецептор СРЧ содержит субъединицу с молекулярной массой 73 кДа, присоединенную N-концом к мембране. Вероятно, рибосома также связывается со специфическими рецепторами, присутствующими в мембране.
3. Рецептор сигнальной последовательности. Сигнальная последовательность на образующейся полипептидной цепи перемещается от СРЧ ко второму рецептору, находящемуся в мембране и называемому рецептором сигнальной последовательности. Об этом свидетельствуют результаты опытов по фотохимическому сшиванию, в которых используется метка, связанная с сигнальной последовательностью препролактина. Предполагаемый мембраносвязанный рецептор представляет собой гликопротеин с молекулярной массой 35 кДа. Возможно, он образует часть канала, через который осуществляется перенос. С помощью такого же подхода с использованием поперечной сшивки и синтетического сигнального пептида был обнаружен еще один кандидат на роль рецептора сигнальной последовательности (45 кДа). Связь между этими двумя белками неизвестна и функции их до конца не установлены. Как только образовавшаяся полипептидная цепь связывается с мембраносвязанным рецепторм, СРЧ и ее рецептор могут освободиться от рибосомы и принять участие в новом цикле. О предполагаемом канале, участвующем в переносе, ничего неизвестно; очистка его является довольно сложной задачей.
Дата: 2019-07-30, просмотров: 346.