Содержание
Введение. 2
1 История химической кинетики. 4
2 Биокинетика. 6
3 Движение клеток. 7
4 Системы подвижности эукариотических клеток. 10
4.1 Микротрубочки. 10
4.1.1 Микротрубочки в ресничках и жгутиках. 11
4.1.2 Самосборка микротрубочек и ее регуляция. 14
4.2 Микрофиламенты.. 15
4.2.1 Свойства актинов немышечных клеток. 17
4.3 Микротрубочки, микрофиламенты и клеточные мембраны.. 18
5 Генерация движения микрофиламентами. 23
5.1 Регуляция скольжения белками микрофиламентов. 25
Список используемой литературы.. 29
Введение
Первый признак, который отличает живое от неживого, это движение, постоянное развитие во времени. Мы заведомо знаем, что события складываются из последовательностей весьма определенных стадий и циклов, разворачиваются во времени. Каждая стадия события имеет продолжительность, определенность и значимость.
Очевидно, что в ритмах живого лежат последовательности превращений молекул. Что определяет протекание биологических процессов во времени? Каковы пути и возможности ускорений биохимических реакций? Какая стадия определяет скорость того или иного биологического явления? Какие события на молекулярном уровне задают динамику развития в целом? Постановка такого рода вопросов связана с развитием области количественных исследований, которая называется биологической кинетикой (биокинетикой).
Исследование количественных закономерностей развития биологических процессов на молекулярном уровне во времени составляет предмет биологической (биохимической) кинетики. В задачи биокинетики входит выяснение механизмов, определяющих скорости и природу процессов.
Изучение динамики биологических процессов охватывает большой круг явлений. Многие из них уже в настоящее время могут быть интерпретированы на молекулярном уровне. За последние десятилетия существенный прогресс в данной области в значительной степени связан с интенсивным изучением ферментов и ферментных систем. Именно ферменты в большинстве случаев являются кинетическими элементами, определяющими скорости и направления развития биопроцессов. Самосогласованность биологических процессов на молекулярном уровне существенным образом определяется отработанными эволюцией процессами обмена информацией с помощью сигнальных молекул и белковых рецепторов. Эти процессы характеризуются вполне определенными кинетическими закономерностями, анализу которых посвящен значительный раздел называемый молекулярной рецепцией.
Наконец, большой и важный раздел современной биокинетики связан с анализом кинетики роста и эволюции клеточных популяций. Клетка как элементарная ячейка жизни представляет собой высокоорганизованный реактор, обладающий свойством полностью воспроизводить себя во всей сложности состава и структуры. Понимание динамики клеточного роста принципиально важно как при решении задач микробиологии, биотехнологии и управляемого биосинтеза, так и для развития количественной медицины, онкологии, для понимания и управления механизмами старения.
Итак, Биокинетика — наука, изучающая на молекулярном уровне закономерности развития биологических процессов в системах in vitro, живых органах и тканях, клеточных популяциях
История химической кинетики
Выделение химической кинетики в самостоятельную научную дисциплину произошло в 80-е гг. XIX в. Это оказалось возможным благодаря накоплению и систематизации громадного фактического материала по изучению механизмов химических превращений.
Вероятно, одной из самых первых попыток изучения механизма химической реакции являются работы Р. Бойля, который в 1680 г. наблюдал яркое свечение фосфора при уменьшении давления воздуха. Несколько позже, в начале – середине XVIII в. А. Лавуазье изучал образование оксида ртути при взаимодействии этого металла с кислородом. В рамках этой работы в 1770 г. Лавуазье провел один из самых длинных за всю историю химии опыт, длившийся 100 дней.
Вслед за Лавуазье ученые-химики не раз предпринимали попытки исследовать закономерности химических превращений, найти материальный субстрат химических реакций
В 1789 г. английский ученый У. Хиггинс предположил, что процесс растворения железа в кислоте является сложной реакцией, протекающей с образованием промежуточных соединений. Эти промежуточные соединения распадаются к моменту окончания реакции из-за действия «силы сродства». Благодаря работе Хиггинса в химию было впервые введено понятие времени протекания химической реакции.
В 1894 г. ученица Хиггинса У. Фульгем показала, что вода способна образовывать промежуточные соединения с реагентами при протекании химических реакций в водных растворах. Фульгем предположила, что вода катализирует реакции, происходящие в растворах.
Гипотезы английских ученых У. Хиггинса и У. Фульгем о наличии промежуточных продуктов химической реакции соответствуют современным представлениям, согласно которым любая химическая реакция представляет собой совокупность элементарных актов (превращений). Эти превращения происходят из-за взаимодействия между собой молекул исходных веществ (субстратов реакции) и приводят к образованию продуктов реакции. При этом любое химическое превращение обратимо.
Впервые предположил обратимый характер химических превращений Д.И. Менделеев в книге «Основы химии» (1869). В этой книге он уделяет особое внимание влиянию внешних условий на закономерности протекания химических реакций и подчеркивает сложный характер химических превращений.
Дальнейшее развитие учения о механизмах химических реакций связано с именами A.M. Бутлерова и М.Д. Львова. В 1861 г. Бутлеров сформулировал теорию химического строения, в которой подчеркивал, что закономерности протекания химических реакций зависят от строения веществ, вступающих в реакцию. Теория химического строения Бутлерова была развита и дополнена его учеником М.Д. Львовым (1884).
Биокинетика
Один из основных разделов кинетики изучает кинетику биологических реакций; этот раздел принято называть биокинетикой. Биокинетика является пограничной наукой, возникшей на стыке биохимии и химической кинетики (см рис. 1). Выделение биокинетики в отдельную дисциплину неслучайно, оно логически оправдано и связано с исключительной значимостью кинетических процессов для всех живых организмов. Биокинетика — относительно молодая наука. Термин «Биокинетика» был введен И.В. Березиным и С.Д. Варфоломеевым в 1979 г. Традиционно в курсе биокинетики рассматриваются ферментативные реакции, процессы взаимодействия лигандов с рецепторами и процессы клеточного роста.

Рисунок 1. « Соотношение между основными естественнонаучными дисциплинами».
Движение клеток
При изучении живых систем часто говорится о движении: движутся хромосомы к полюсам клетки во время митоза, перемещаются вакуоли клеточных органелл, движется клеточная поверхность. Кроме того, в клетках растений и животных наблюдаются токи цитоплазмы (например, в растительных клетках или у амебы). Более того, отдельные клетки (свободноживущие одноклеточные организмы или специфические типы клеток в многоклеточных животных организмах) обладают способностью активно перемещаться, «ползать» (см. рис. 3.). Некоторые клетки имеют специализированные структуры, реснички или жгутики, которые позволяют им или самым перемещаться, или перемещать окружающую их жидкость. Наконец, у многоклеточных животных организмов есть специализированные клетки, мышечная работа которых позволяет производить различные движения органов, отдельных его частей и всего организма. В основе всех этих многочисленных двигательных реакций лежат общие молекулярные механизмы. Кроме того, наличие каких-либо двигательных аппаратов должно сочетаться и структурно связываться с существованием опорных, каркасных или скелетных внутриклеточных образований. Поэтому можно говорить (описывать и изучать) об опорно-двигательной системе клеток.
Само понятие о цитоскелете или скелетных компонентах цитоплазмы разных клеток было высказано Н.К.Кольцовым, выдающимся русским цитологом еще в начале XX века. К сожалению, они были забыты и только в конце 1950-х годов с помощью электронного микроскопа эта скелетная система была переоткрыта.
Огромный вклад в изучение цитоскелета внес метод иммунофлуоресценции, который помог разобраться в химии и динамике этого чрезвычайно важного компонента клетки. Цитоскелетные компоненты представлены нитевидными, неветвящимися белковыми комплексами, или филаментами (тонкими нитями).
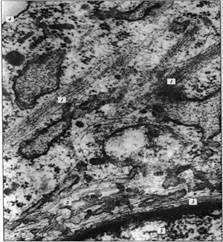
Рисунок 2. «Микрофотография элементов цитоскелета, полученная с помощью электронного микроскопа». 1 - пучки микрофиламентов; 2 - микротрубочки; 3 - промежуточные филаменты 4 плазматическая мембрана; 5 – ядро.
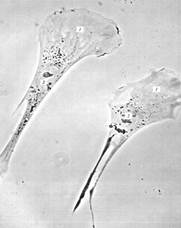
Рисунок 3. «Поляризованные движущиеся фибробласты в культуре ткани». 1 – ламеллоплазма; 2 — ядро
Существуют три системы филаментов, различающихся по химическому составу, ультраструктуре и функциональным свойствам. Самые тонкие нити — это микрофиламенты; их диаметр составляет около 6 мм, и состоят они в основном из белка актина. К другой группе нитчатых структур относятся микротрубочки, которые имеют диаметр 25 нм и состоят в основном из белка тубулина. Третья группа представлена промежуточными филаментами с диметром около 10 нм (промежуточным по сравнению с 6 и 25 нм), образующимися из разных, но родственных белков (рис. 2 и 4).
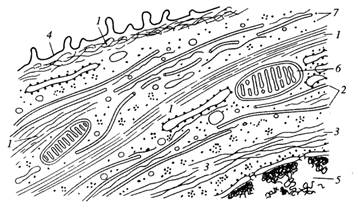
Рисунок 4. «Схематическое изображение цитоскелетных компонентов клеток». 1 – микрофиламенты; 2 – микротрубочки; 3 – промежуточные филаменты; 4 – плазмалитическая мембрана; 5 – ядро; 6 – митохондрии; 7 – рибосомы.
Микротрубочки
Микротрубочки предстали как особые внутриклеточные структуры (особенно внутри жгутика) благодаря применению методов электронной микроскопии. В 1946 г. Джакус и Холл продемонстрировали наличие одинаковых по диаметру трубчатых структур в ресничках Paramecium. Впоследствии трубчатые структуры обнаружили почти во всех клетках. Стали считать, что они характерны для эукариотичсского уровня организации. Однако микротубулярные структуры были обнаружены и у прокариот — в цитоплазме спирохет. Морфологически и биохимически эти микротрубочки подобны тем, которые наблюдаются в эукариотических клетках. Это открытие поддержало «экзогенную гипотезу» происхождения микротрубочек эукариот, согласно которой реснички и жгутики клеток высших организмов считаются приобретенными извне—путем симбиоза клеток, ранее не имевших жгутиков или ресничек, со спирохетами, содержавшими микротрубочки.
Микротрубочки представляют собой длинные полые цилиндры, наружный диаметр которых около 24 нм, а внутренний—15 нм. В большинстве клеток длина трубочек обычно не превышает нескольких микрон, хотя в некоторых специализированных клетках, например в моторных нейронах центральной нервной системы, трубочки могут быть длиной в несколько сантиметров. Стенка микротрубочки (около 5 нм толщиной) построена из продольно ориентированных протофибрилл. Протофибриллы состоят из глобулярных субъединиц (их очень хорошо видно в электронный микроскоп при негативном окрашивании), содержащих только один белок – тубулин (рис. 5). Помимо того, что микротрубочки имеют прямое отношение к подвижности клетки, они участвуют и в других процессах, более или менее связанных с подвижностью, например в поддержании формы клетки, во внутриклеточном транспорте веществ, в секреции клеточных продуктов, в движении хромосом при делении клетки и, возможно, в осуществлении сенсорных связей, а также в перемещении компонентов клеточной мембраны. Микротрубочки могут быть рассеяны по всей цитоплазме, а могут быть собраны в организованные структуры.
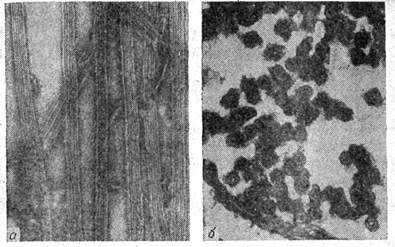
Рисунок 5. «Электронные, микрофотографии микротрубочек в краевых областях эритроцитов тритона»: а — продольный срез, на котором: видна линейная упаковка субъединиц в микротрубочках; б—поперечный срез; видно, что стенка каждой микротрубочки образована тринадцатью субъединицами. Негативное окрашивание. ×200 000.
Чтобы понять, как клетка движется с помощью микротрубочек, мы рассмотрим их организацию в ресничках жгутиках эукариотических клеток, в сократимом аксостиле некоторых жгутиконосцев, а также в микротубулярной сети цитоплазмы животных клеток.
Микрофиламенты
Микрофиламенты эукариотических клеток представляют собой длинные нитевидные структуры толщиной 5-7 нм, находящиеся в цитоплазме. Они состоят главным образом из актина (хотя можно обнаружить и другие белки), в связи, с чем их часто называют актиновыми филаментами (или нитями). Они иногда образуют группы или пучки, а в некоторых высоко специализированных клетках, например в мышечных волокнах, упорядоченные и стабильные структуры. Чаще, однако из микрофиламентов формируются нестабильные пучки или тонкие сетчатые структуры, их форма и локализация в клетке изменяются в зависимости от фазы жизненного цикла клетки, ее движения и т. п. Это означает, что, подобно цитоплазматическим микротрубочкам, микрофиламенты немышечных клеток представляют собой изменяемые структуры, способные к самосборке или деполимеризации на составляющие их молекулы в зависимости от нужд клетки.
Морфологию микрофиламентов и их поведение в процессе клеточного движения и цикла развития клетки много изучали на культуре животных клеток с помощью флуоресцирующих антител против актина, а также против других белков (миозин, тропомиозин и а-актинин), которые часто обнаруживаются в микрофиламентах. Когда клетки находятся в состоянии покоя и прикреплены к стенкам сосуда, в котором живут, в них можно наблюдать длинные пучки микрофиламентов, тянущиеся сквозь всю клетку, по-видимому, непосредственно под клеточной мембраной. В «возбужденные» выступы клетки с нестабильными, как бы волнующимися краями они обычно не заходят. Пучки микрофиламентов называются стрессовыми, типичны для нормальных клеток и отсутствуют в клетках, трансформированных онкогенными вирусами. Когда клетки движутся, в дополнение к стрессовым волокнам появляются топкие пучки актиновых нитей, рассеянные в цитоплазме; они особенно заметны в направленных вперед выступах клеток.
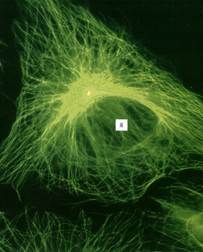
Рисунок 9. «Сеть микротрубочек, окрашенная мечеными антителами к тубулину в клетке культуры ткани в Gj-периоде». В клеточном центре желтым окрашена центриоль, связанная с антителами к γ-тубулину, Я – ядро.
Список используемой литературы
1.Биологический энциклопедический словарь. / Гл. ред. М. С. Гилярон; Редкол.: А. А. Баев, Г. Г. Винберг, Г. А. Заварзип и др. — 2-е изд., исправл. — М.: Сов. энциклопедия, 1989. — 864 с, ил.
2.Варфоломеев С. Д., Гуревич К. Г. Биокинетика: Практический курс. – М.: ФАИР-ПРЕСС, 1999.– 720 с: ил.
3.Мецлер Д. Биохимия: Химические реакции в живой клетке. В 3-х томах том 1, 2, 3. Пер. с англ. – М.: Изд-во «Мир», – 1980.
4.Ченцов Ю.С. Введение в клеточную биологию: Учебник для вузов. – 4-е изд., перераб, и доп. / Ю.С. Ченцов. – М.: ИКЦ «Академкнига», 2004. – 495 с: ил.
5.Юрина Н. А., Радостина А. И. Гистология: Учебник. – М.: Медицина, 1995. – 256 с; ил.
6.Гистология: Учебник. 2-е изд., перераб, и доп. / Под ред. Э.Г. Улумбекова, Ю./ Челышева. – М.: ГЭОТАР-МЕД, 2002. – 672 с: ил.
Содержание
Введение. 2
1 История химической кинетики. 4
2 Биокинетика. 6
3 Движение клеток. 7
4 Системы подвижности эукариотических клеток. 10
4.1 Микротрубочки. 10
4.1.1 Микротрубочки в ресничках и жгутиках. 11
4.1.2 Самосборка микротрубочек и ее регуляция. 14
4.2 Микрофиламенты.. 15
4.2.1 Свойства актинов немышечных клеток. 17
4.3 Микротрубочки, микрофиламенты и клеточные мембраны.. 18
5 Генерация движения микрофиламентами. 23
5.1 Регуляция скольжения белками микрофиламентов. 25
Список используемой литературы.. 29
Введение
Первый признак, который отличает живое от неживого, это движение, постоянное развитие во времени. Мы заведомо знаем, что события складываются из последовательностей весьма определенных стадий и циклов, разворачиваются во времени. Каждая стадия события имеет продолжительность, определенность и значимость.
Очевидно, что в ритмах живого лежат последовательности превращений молекул. Что определяет протекание биологических процессов во времени? Каковы пути и возможности ускорений биохимических реакций? Какая стадия определяет скорость того или иного биологического явления? Какие события на молекулярном уровне задают динамику развития в целом? Постановка такого рода вопросов связана с развитием области количественных исследований, которая называется биологической кинетикой (биокинетикой).
Исследование количественных закономерностей развития биологических процессов на молекулярном уровне во времени составляет предмет биологической (биохимической) кинетики. В задачи биокинетики входит выяснение механизмов, определяющих скорости и природу процессов.
Изучение динамики биологических процессов охватывает большой круг явлений. Многие из них уже в настоящее время могут быть интерпретированы на молекулярном уровне. За последние десятилетия существенный прогресс в данной области в значительной степени связан с интенсивным изучением ферментов и ферментных систем. Именно ферменты в большинстве случаев являются кинетическими элементами, определяющими скорости и направления развития биопроцессов. Самосогласованность биологических процессов на молекулярном уровне существенным образом определяется отработанными эволюцией процессами обмена информацией с помощью сигнальных молекул и белковых рецепторов. Эти процессы характеризуются вполне определенными кинетическими закономерностями, анализу которых посвящен значительный раздел называемый молекулярной рецепцией.
Наконец, большой и важный раздел современной биокинетики связан с анализом кинетики роста и эволюции клеточных популяций. Клетка как элементарная ячейка жизни представляет собой высокоорганизованный реактор, обладающий свойством полностью воспроизводить себя во всей сложности состава и структуры. Понимание динамики клеточного роста принципиально важно как при решении задач микробиологии, биотехнологии и управляемого биосинтеза, так и для развития количественной медицины, онкологии, для понимания и управления механизмами старения.
Итак, Биокинетика — наука, изучающая на молекулярном уровне закономерности развития биологических процессов в системах in vitro, живых органах и тканях, клеточных популяциях
История химической кинетики
Выделение химической кинетики в самостоятельную научную дисциплину произошло в 80-е гг. XIX в. Это оказалось возможным благодаря накоплению и систематизации громадного фактического материала по изучению механизмов химических превращений.
Вероятно, одной из самых первых попыток изучения механизма химической реакции являются работы Р. Бойля, который в 1680 г. наблюдал яркое свечение фосфора при уменьшении давления воздуха. Несколько позже, в начале – середине XVIII в. А. Лавуазье изучал образование оксида ртути при взаимодействии этого металла с кислородом. В рамках этой работы в 1770 г. Лавуазье провел один из самых длинных за всю историю химии опыт, длившийся 100 дней.
Вслед за Лавуазье ученые-химики не раз предпринимали попытки исследовать закономерности химических превращений, найти материальный субстрат химических реакций
В 1789 г. английский ученый У. Хиггинс предположил, что процесс растворения железа в кислоте является сложной реакцией, протекающей с образованием промежуточных соединений. Эти промежуточные соединения распадаются к моменту окончания реакции из-за действия «силы сродства». Благодаря работе Хиггинса в химию было впервые введено понятие времени протекания химической реакции.
В 1894 г. ученица Хиггинса У. Фульгем показала, что вода способна образовывать промежуточные соединения с реагентами при протекании химических реакций в водных растворах. Фульгем предположила, что вода катализирует реакции, происходящие в растворах.
Гипотезы английских ученых У. Хиггинса и У. Фульгем о наличии промежуточных продуктов химической реакции соответствуют современным представлениям, согласно которым любая химическая реакция представляет собой совокупность элементарных актов (превращений). Эти превращения происходят из-за взаимодействия между собой молекул исходных веществ (субстратов реакции) и приводят к образованию продуктов реакции. При этом любое химическое превращение обратимо.
Впервые предположил обратимый характер химических превращений Д.И. Менделеев в книге «Основы химии» (1869). В этой книге он уделяет особое внимание влиянию внешних условий на закономерности протекания химических реакций и подчеркивает сложный характер химических превращений.
Дальнейшее развитие учения о механизмах химических реакций связано с именами A.M. Бутлерова и М.Д. Львова. В 1861 г. Бутлеров сформулировал теорию химического строения, в которой подчеркивал, что закономерности протекания химических реакций зависят от строения веществ, вступающих в реакцию. Теория химического строения Бутлерова была развита и дополнена его учеником М.Д. Львовым (1884).
Биокинетика
Один из основных разделов кинетики изучает кинетику биологических реакций; этот раздел принято называть биокинетикой. Биокинетика является пограничной наукой, возникшей на стыке биохимии и химической кинетики (см рис. 1). Выделение биокинетики в отдельную дисциплину неслучайно, оно логически оправдано и связано с исключительной значимостью кинетических процессов для всех живых организмов. Биокинетика — относительно молодая наука. Термин «Биокинетика» был введен И.В. Березиным и С.Д. Варфоломеевым в 1979 г. Традиционно в курсе биокинетики рассматриваются ферментативные реакции, процессы взаимодействия лигандов с рецепторами и процессы клеточного роста.
Рисунок 1. « Соотношение между основными естественнонаучными дисциплинами».
Движение клеток
При изучении живых систем часто говорится о движении: движутся хромосомы к полюсам клетки во время митоза, перемещаются вакуоли клеточных органелл, движется клеточная поверхность. Кроме того, в клетках растений и животных наблюдаются токи цитоплазмы (например, в растительных клетках или у амебы). Более того, отдельные клетки (свободноживущие одноклеточные организмы или специфические типы клеток в многоклеточных животных организмах) обладают способностью активно перемещаться, «ползать» (см. рис. 3.). Некоторые клетки имеют специализированные структуры, реснички или жгутики, которые позволяют им или самым перемещаться, или перемещать окружающую их жидкость. Наконец, у многоклеточных животных организмов есть специализированные клетки, мышечная работа которых позволяет производить различные движения органов, отдельных его частей и всего организма. В основе всех этих многочисленных двигательных реакций лежат общие молекулярные механизмы. Кроме того, наличие каких-либо двигательных аппаратов должно сочетаться и структурно связываться с существованием опорных, каркасных или скелетных внутриклеточных образований. Поэтому можно говорить (описывать и изучать) об опорно-двигательной системе клеток.
Само понятие о цитоскелете или скелетных компонентах цитоплазмы разных клеток было высказано Н.К.Кольцовым, выдающимся русским цитологом еще в начале XX века. К сожалению, они были забыты и только в конце 1950-х годов с помощью электронного микроскопа эта скелетная система была переоткрыта.
Огромный вклад в изучение цитоскелета внес метод иммунофлуоресценции, который помог разобраться в химии и динамике этого чрезвычайно важного компонента клетки. Цитоскелетные компоненты представлены нитевидными, неветвящимися белковыми комплексами, или филаментами (тонкими нитями).
Рисунок 2. «Микрофотография элементов цитоскелета, полученная с помощью электронного микроскопа». 1 - пучки микрофиламентов; 2 - микротрубочки; 3 - промежуточные филаменты 4 плазматическая мембрана; 5 – ядро.
Рисунок 3. «Поляризованные движущиеся фибробласты в культуре ткани». 1 – ламеллоплазма; 2 — ядро
Существуют три системы филаментов, различающихся по химическому составу, ультраструктуре и функциональным свойствам. Самые тонкие нити — это микрофиламенты; их диаметр составляет около 6 мм, и состоят они в основном из белка актина. К другой группе нитчатых структур относятся микротрубочки, которые имеют диаметр 25 нм и состоят в основном из белка тубулина. Третья группа представлена промежуточными филаментами с диметром около 10 нм (промежуточным по сравнению с 6 и 25 нм), образующимися из разных, но родственных белков (рис. 2 и 4).
Рисунок 4. «Схематическое изображение цитоскелетных компонентов клеток». 1 – микрофиламенты; 2 – микротрубочки; 3 – промежуточные филаменты; 4 – плазмалитическая мембрана; 5 – ядро; 6 – митохондрии; 7 – рибосомы.
Системы подвижности эукариотических клеток
Микротрубочки
Микротрубочки предстали как особые внутриклеточные структуры (особенно внутри жгутика) благодаря применению методов электронной микроскопии. В 1946 г. Джакус и Холл продемонстрировали наличие одинаковых по диаметру трубчатых структур в ресничках Paramecium. Впоследствии трубчатые структуры обнаружили почти во всех клетках. Стали считать, что они характерны для эукариотичсского уровня организации. Однако микротубулярные структуры были обнаружены и у прокариот — в цитоплазме спирохет. Морфологически и биохимически эти микротрубочки подобны тем, которые наблюдаются в эукариотических клетках. Это открытие поддержало «экзогенную гипотезу» происхождения микротрубочек эукариот, согласно которой реснички и жгутики клеток высших организмов считаются приобретенными извне—путем симбиоза клеток, ранее не имевших жгутиков или ресничек, со спирохетами, содержавшими микротрубочки.
Микротрубочки представляют собой длинные полые цилиндры, наружный диаметр которых около 24 нм, а внутренний—15 нм. В большинстве клеток длина трубочек обычно не превышает нескольких микрон, хотя в некоторых специализированных клетках, например в моторных нейронах центральной нервной системы, трубочки могут быть длиной в несколько сантиметров. Стенка микротрубочки (около 5 нм толщиной) построена из продольно ориентированных протофибрилл. Протофибриллы состоят из глобулярных субъединиц (их очень хорошо видно в электронный микроскоп при негативном окрашивании), содержащих только один белок – тубулин (рис. 5). Помимо того, что микротрубочки имеют прямое отношение к подвижности клетки, они участвуют и в других процессах, более или менее связанных с подвижностью, например в поддержании формы клетки, во внутриклеточном транспорте веществ, в секреции клеточных продуктов, в движении хромосом при делении клетки и, возможно, в осуществлении сенсорных связей, а также в перемещении компонентов клеточной мембраны. Микротрубочки могут быть рассеяны по всей цитоплазме, а могут быть собраны в организованные структуры.
Рисунок 5. «Электронные, микрофотографии микротрубочек в краевых областях эритроцитов тритона»: а — продольный срез, на котором: видна линейная упаковка субъединиц в микротрубочках; б—поперечный срез; видно, что стенка каждой микротрубочки образована тринадцатью субъединицами. Негативное окрашивание. ×200 000.
Чтобы понять, как клетка движется с помощью микротрубочек, мы рассмотрим их организацию в ресничках жгутиках эукариотических клеток, в сократимом аксостиле некоторых жгутиконосцев, а также в микротубулярной сети цитоплазмы животных клеток.
Дата: 2019-07-24, просмотров: 521.