Газовая смесь, поступившая в легкие при вдохе, распределяется на две части. Одна из них не принимает участие в газообмене, т.к. заполняет воздухоносные пути (анатомически мертвое пространство). Другая часть (альвеолярный объем) поступает в респираторный отдел (альвеолярные протоки, мешочки и альвеолы), где принимает участие в газообмене. Она обеспечивает вентиляцию альвеолярного пространства.
Особенности альвеолярной вентиляции:
-интенсивность обновления газового состава, определяемая соотношением альвеолярного объема и альвеолярной вентиляции.
-изменения альвеолярного объема (увеличение/уменьшение размера вентилируемых альвеол, либо изменение кол-ва альвеол, участвующих в вентиляции).
-различия внутрилегочных характеристик сопротивления и эластичности, приводящее к асинхронности альвеолярной вентиляции.
-поток газов в альвеолу или из нее определяется механическими характеристиками легких и дыхательных путей, а также силами (или давлением), воздействующими на них. Механические характеристики обусловлены сопротивлением дыхательных путей потоку воздуха и эластическими св-ва легочной паренхимы.
Размеры альвеолярного пространства таковы, что смешивание газов в альвеолярной единице происходит практически мгновенно как следствие дыхательных движений, кровотока и диффузии.
Неравномерность альвеолярной вентиляции обусловлена и гравитационным фактором-разницей транспульмонального давления в верхних и нижних отделах грудной клетки. В вертикальном положении в нижних отделах это давление выше примерно на 8 см.вод.ст. Апико-базальный градиент всегда присутствует независимо от степени наполнения легких→определяет наполнение воздухом альвеол в разных отделах легких.
В норме вдыхаемый газ смешивается мгновенно с альвеолярным газом. Состав газа в альвеолах практически гомогенен в любую респираторную фазу и в любой момент вентиляции.
Любое повышение альвеолярного транспорта кислорода и углекислого газа (напр. при физических нагрузках) сопровождается повышение градиентов концентрации газов, которые способствуют возрастанию их смешивания в альвеолах. Нагрузка стимулирует альвеолярное смешивание путем повышения потока вдыхаемого воздуха и возрастания кровотока, повышает альвеолярно-капиллярный градиент давления для кислорода и углекислого газа.
Диффузия газов.
Газообмен — совокупность процессов, обеспечивающих переход кислорода внешней среды в ткани живого организма, а углекислого газа из тканей во внешнюю среду.
Перемещение газов осуществляется под влиянием разности парциальных давлений и напряжений этих газов в каждой из сред организма.
Парциальное давление кислорода в воздухе, заполняющем альвеолы легких, около 100 мм рт. ст., а его напряжение в венозной крови, притекающей к легким, около 40 мм рт. ст. Вследствие разности давлений кислород из альвеол направляется в кровь, где связывается с гемоглобином эритроцитов. Парциальное давление углекислого газа в альвеолярном воздухе составляет 40 мм рт. ст., а его напряжение в притекающей к легким венозной крови — 48 мм рт. ст. Вследствие разности давлений углекислый газ переходит в альвеолы.
В артериальной крови, притекающей к тканям, напряжение кислорода выше, чем в тканях, а напряжение углекислого газа, наоборот, значительно ниже. Вследствие этого кислород переходит из крови в ткани и включается в цикл метаболических процессов, а углекислый газ, в избытке содержащийся в тканях, переходит в кровь и переносится затем в лёгкие. Процесс газообмена происходит непрерывно до тех пор, пока существует разность парциальных давлений и напряжений газов в каждой из сред, участвующих в газообмене решающим фактором, обусловливающим непрерывность газообмена, является постоянство газового состава альвеолярного воздуха.
Величина газообмена является показателем интенсивности окислительных процессов, протекающих в тканях. Об уровне газообмена можно судить и по величине минутной вентиляции легких. При спокойном дыхании через легкие проходит около 8000 мл воздуха в 1 мин. При физических и эмоциональных напряжениях, различных заболеваниях, сопровождающихся усилением окислительных процессов в тканях, легочная вентиляция возрастает.
Вентиляционно-перфузионные отношения в разных отделах легкого.
Кровоток в капиллярах легких и легочная вентиляция неодинаковы в различных отделах и зависят от положения тела.
Основное влияние на распределение перфузии в легких оказывает гравитация, что обусловлено низким АД в системе малого круга кровообращения (15-20 мм рт. ст.). Поэтому при любом положении тела в пространстве нижние отделы легких по сравнению с верхними будут иметь больший кровоток.
Зависимость перфузии от сил гравитации более выражена, чем у вентиляции, что определяет и характер изменения вентиляционно-перфузионных отношений по направлению от верхушек к основанию легких. Нормальная альвеолярная вентиляция (VA) у взрослых составляет ~ 4 л/мин, а общая легочная перфузия (Q) ~ 5 л/мин. Следовательно, отношение величин вентиляции и перфузии будет равно 4/5, или 0,8. Изменение отношения YA /Q будет отражать степень гипервентиляции (гипоперфузии) или гиперперфузии (гиповентиляции) в целом легком или в его отдельных зонах.
Распределение вентиляции зависит от нескольких факторов. Основным является растяжимость легочной ткани, которая неодинакова в различных легочных зонах.
Транспорт кислорода.
Транспорт О2 осуществляется в физически растворенном и химически связанном виде. Физические процессы, т. е. растворение газа, не могут обеспечить запросы организма в О2.
Согласно закону Фика, газообмен О2 между альвеолярным воздухом и кровью происходит благодаря наличию концентрационного градиента О2 между этими средами. В альвеолах легких парциальное давление О2 составляет 13,3 кПа, или 100 мм рт.ст., а в притекающей к легким венозной крови парциальное напряжение О2 составляет примерно 5,3 кПа, или 40 мм рт.ст. Давление газов в воде или в тканях организма обозначают термином «напряжение газов» и обозначают символами Ро2, Рсo2.
Транспорт О2 начинается в капиллярах легких после его химического связывания с гемоглобином. Гемоглобин (Нb) способен избирательно связывать О2 и образовывать оксигемоглобин (НbО2) в зоне высокой концентрации О2 в легких и освобождать молекулярный О2 в области пониженного содержания О2 в тканях. При этом свойства гемоглобина не изменяются и он может выполнять свою функцию на протяжении длительного времени.
Гемоглобин переносит О2 от легких к тканям. Эта функция зависит от двух свойств гемоглобина: 1) способности изменяться от восстановленной формы, которая называется дезоксигемоглобином, до окисленной (Нb + О2 à НbО2) с высокой скоростью (полупериод 0,01 с и менее) при нормальном Рог в альвеолярном воздухе; 2) способности отдавать О2 в тканях (НbО2 à Нb + О2) в зависимости от метаболических потребностей клеток организма.
Зависимость степени оксигенации гемоглобина от парциального давления Ог в альвеолярном воздухе графически представляется в виде кривой диссоциации оксигемоглобина, или сатурационной кривой. Плато кривой диссоциации характерно для насыщенной О2 (сатурированной) артериальной крови, а крутая нисходящая часть кривой — венозной, или десатурированной, крови в тканях.
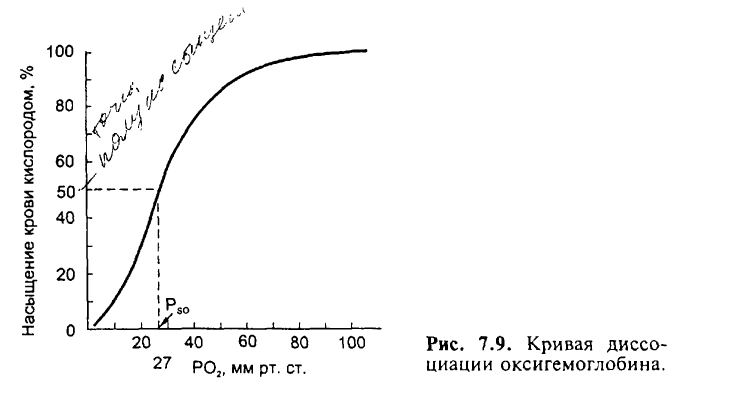
На сродство кислорода к гемоглобину влияют различные метаболические факторы, что выражается в виде смещения кривой диссоциации влево или вправо. Сродство гемоглобина к кислороду регулируется важнейшими факторами метаболизма тканей: Ро2 pH, температурой и внутриклеточной концентрацией 2,3-дифосфоглицерата. Величина рН и содержание СО2 в любой части организма закономерно изменяют сродство гемоглобина к О2: уменьшение рН крови вызывает сдвиг кривой диссоциации соответственно вправо (уменьшается сродство гемоглобина к О2), а увеличение рН крови — сдвиг кривой диссоциации влево (повышается сродство гемоглобина к О2). Например, рН в эритроцитах на 0,2 единицы ниже, чем в плазме крови. В тканях вследствие повышенного содержания СО2 рН также меньше, чем в плазме крови. Влияние рН на кривую диссоциации оксигемоглобина называется «эффектом Бора».
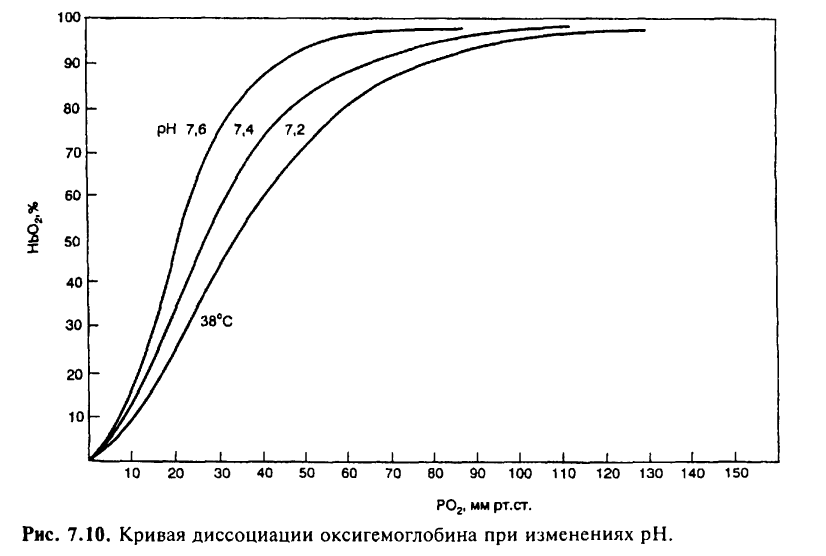
Рост температуры уменьшает сродство гемоглобина к О2. В работающих мышцах увеличение температуры способствует освобождению О2. Уменьшение температуры тканей или содержания 2,3-дифосфоглицерата вызывает сдвиг влево кривой диссоциации оксигемоглобина.
Метаболические факторы являются основными регуляторами связывания О2 с гемоглобином в капиллярах легких, когда уровень O2, рН и СО2 в крови повышает сродство гемоглобина к О2 по ходу легочных капилляров. В условиях тканей организма эти же факторы метаболизма понижают сродство гемоглобина к О2 и способствуют переходу оксигемоглобина в его восстановленную форму — дезоксигемоглобин. В результате О2 по концентрационному градиенту поступает из крови тканевых капилляров в ткани организма.
Оксид углерода (II) — СО, способен соединяться с атомом железа гемоглобина, изменяя его свойства и реакцию с О2. Очень высокое сродство СО к Нb (в 200 раз выше, чем у О2) блокируют один или более атомов железа в молекуле гема, изменяя сродство Нb к О2.
Под кислородной емкостью крови понимают количество Ог, которое связывается кровью до полного насыщения гемоглобина. При содержании гемоглобина в крови 8,7 ммоль*л-1 кислородная емкость крови составляет 0,19 мл О2 в 1 мл крови (температура 0oC и барометрическое давление 760 мм рт.ст., или 101,3 кПа). Величину кислородной емкости крови определяет количество гемоглобина, 1 г которого связывает 1,36—1,34 мл О2. Кровь человека содержит около 700—800 г гемоглобина и может связать таким образом почти 1 л О2. Физически растворенного в 1 мл плазмы крови О2 очень мало (около 0,003 мл), что не может обеспечить кислородный запрос тканей. Растворимость О2 в плазме крови равна 0,225 мл*л-1*кПа-1.
Обмен О2 между кровью капилляров и клетками тканей также осуществляется путем диффузии. Концентрационный градиент О2 между артериальной кровью (100 мм рт.ст., или 13,3 кПа) и тканями (около 40 мм рт.ст., или 5,3 кПа) равен в среднем 60 мм рт.ст. (8,0 кПа). Изменение градиента может быть обусловлено как содержанием О2 в артериальной крови, так и коэффициентом утилизации О2, который составляет в среднем для организма 30— 40%. Коэффициентом утилизации кислорода называется количество О2, отданного при прохождении крови через тканевые капилляры, отнесенное к кислородной емкости крови.
С другой стороны, известно, что при напряжении О2 в артериальной крови капилляров, равном 100 мм рт.ст. (13,3 кПа), на мембранах клеток, находящихся между капиллярами, эта величина не превышает 20 мм рт.ст. (2,7 кПа), а в митохондриях равна в среднем 0,5 мм рт.ст. (0,06 кПа).
Дата: 2019-04-23, просмотров: 373.