Основные физиологические свойства сердечной мышцы. Как всякая мышца, сердечная обладает возбудимостью, сократимостью и проводимостью. Кроме того, сердце обладает еще и способностью к автоматии. При действии раздражителя на миокард возникает его возбуждение и сокращение. При этом на пороговую силу сердце отвечает максимальным сокращением (закон "все или ничего" Боудича). Однако, сердце не при всех условиях отвечает на раздражение одинаково. Величина "все" может меняться в зависимости от температуры, растяжения мышцы, степени ее утомления, состава протекающей крови и т.п.
В состоянии покоя поверхностная мембрана мышечной клетки поляризована, и мембранный потенциал равен 80-90 мВ. При возбуждении вследствие перехода катионов Na и Са через мембрану внутрь волокна возникает ПД, равный 100-120 мВ. После быстрой деполяризации наступает несравнимо более медленная реполяризация. Сначала разность потенциалов быстро падает, потом некоторое время удерживается на одном уровне (плато), после чего происходит относительно быстрое восстановление мембранного потенциала до исходной величины.
Описанные изменения электрического состояния мембраны характерны для мышечных волокон желудочков сердца. В других мышечных волокнах сердца форма ПД может быть несколько другой. Наибольшие различия отмечаются в пейсмекерах (водителях ритма), в которых происходит спонтанная медленная деполяризация во время диастолы, а пик и плато в достаточной мере сглажены, хотя и продолжаются то же время. ПД в миокарде длится дольше, чем в волокнах скелетной мускулатуры. Общая его продолжительность равна 0,3 сек при ритме работы 70 в мин. Длительность ПД укорачивается при учащении и удлиняется при замедлении работы сердца.
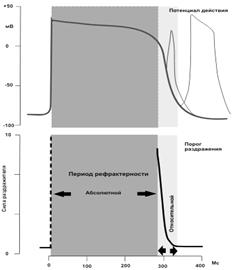
Рис. 19 Типичный вид потенциала действия в миокарде и периоды абсолютной и относительной рефрактерности в цикле возбуждения миокардиоцита
Во время возбуждения сердечная мышца, так же, как и скелетная, утрачивает способность отвечать второй вспышкой возбуждения на искусственное раздражение или на приходящий к ней импульс от очага автоматии. Длительность периода абсолютной рефрактерности в сердце велика и равна 0,27 сек при ритме 70 в мин. Период рефрактерности сердечной мышцы продолжается столько же времени, сколько длится ее сокращение в ответ на одиночное раздражение. Поэтому сердце не способно сокращаться тетанически. По окончании абсолютного рефрактерного периода возбудимость постепенно восстанавливается до исходного уровня. Это - период относительной рефрактерности, который длится 0,03 сек. За ним наступает короткий период супернормальной возбудимости.
Сократимость сердечной мышцы. Процесс сопряжения возбуждения и сокращения в сердце - сложный процесс, происходящий при участии саркоплазматического ретикулюма и ионов Са++. В сердечных волокнах Т-системы выражены слабее, чем в скелетных мышцах, особенно - в предсердиях. Происходит это потому, что волокна сердечной мышцы тоньше, чем скелетной, и саркоплазматический ретикулюм расположен близко к поверхностной мембране. Поэтому в сердце большую роль играет внеклеточный Са++. Это доказывается опытами с перфузией изолированного сердца растворами, не содержащими Са++. В этом случае можно зарегистрировать ПД на мембране клетки, а сокращения мышцы не происходит. Период сокращения отдельных мышечных волокон сердца примерно соответствует длительности ПД. При увеличении частоты возникающих в сердце импульсов укорачивается и продолжительность ПД, и длительность сокращения.
Непосредственным поставщиком энергии для сокращения сердца являются АТФ и КФ. Ресинтез этих соединений происходит за счет энергии дыхательного и гликолитического фосфорилирования. При этом углевод (глюкоза) не являются единственными или преобладающими веществами, используемыми сердечной мышцей для обмена. Только 15% энергии получает сердце за счет глюкозы. Остальное - за счет молочной кислоты, жирных кислот и т.п., т.е. отбросов метаболизма других органов. В сердечной мышце преобладают аэробные процессы.
16-3. Автоматия сердца: механизм, градиент автоматии и его доказательство. Экстрасистолы: понятие, виды, причины их возникновения.
Возникновение и распространение возбуждения. Ритмические сокращения сердца возникают под действием импульсов, возникающих в нем самом, а характерным свойством сердца является автоматия – способность сокращаться под действием собственных импульсов. Автоматию легко наблюдать на выделенном сердце лягушки, помещенном в раствор Рингера, в нем сердце может циклы сокращение-расслабление в течение десятков минут.
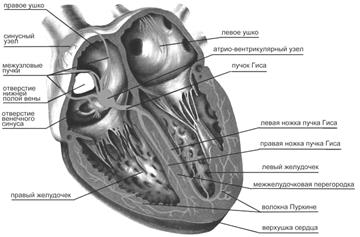
Рис. 20. Проводящая система сердца человека. Подробнее в тексте.
Ритмические импульсы генерируются специальными клетками пейсмекера и распространяются по проводящей системе. В норме водителем ритма служит синоатриальный узел (СА), расположенный в стенке правого предсердия в месте впадения в него верхней полой вены (рис. 20). Частота электрических разрядов СА в покое составляет около 70 в мин. От СА возбуждение вначале распространяется к рабочему миокарду предсердий. Атриовентрикулярная перегородка является электрически невозбудимым образованием, поэтому к желудочкам возбуждение может пройти только единственным путем – по проводящей системе. Из СА возбуждение передается в атриовентрикулярный узел (АВ). После него переходит в пучок Гиса и его левую и правую ножки, заканчиваясь в конечных разветвлениях – волокнах Пуркинье. По проводящей системе возбуждение распространяется со скоростью до 4 м/с. Для сравнения – по рабочему миокарду возбуждение движется со скоростью всего около 0,5 м/с. Высокая скорость проведения импульса по проводящей системе желудочков обеспечивает их синхронное возбуждение. Это повышает эффективность насосной функции сердца. Например, повреждение пучка Гиса может привести к потере 50% мощности сокращений миокарда из-за увеличения длительности асинхронного сокращения.
От ведущего узла по предсердиям возбуждение проводится по специальным пучкам, образованным атипическими мышечными волокнами. Кисом и Флеком описан также пучок атипических волокон, соединяющий синусный узел с атриовентрикулярным узлом (Ашоф-Товара), расположенным в правом предсердии, в области межпредсердной перегородки, вблизи от соединительно-тканного кольца, отделяющего предсердие от желудочка. От атриовентрикулярного узла берет начало пучок Гисса, который, войдя в желудочек по межжелудочковой перегородке, делится на две ветви - правую и левую ножки пучка Гисса. Конечные разветвления проводящей системы представлены широко распространенной, расположенной под эндокардом сетью волокон Пуркинье, которые через т.н. транзиторные клетки соединяются с мышечными волокнами сердца (сократительным миокардом). По разветвлениям проводящей системы возбуждение доходит до всей массы сердечной мышцы, вызывая ее сокращение.
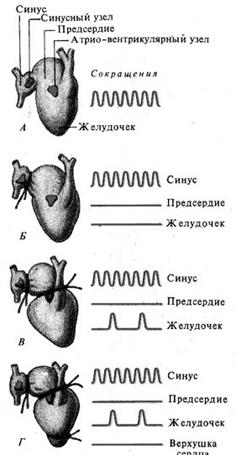 Соподчинение водителей ритма. Все отделы проводящей системы способны спонтанно генерировать импульсы, но с различной частотой. Самой большой частотой обладают клетки СА. Все другие имеют существенно меньшую частоту разрядов, причем, чем дальше отдел проводящей системы расположен от СА, тем его частота ниже. Благодаря этому в норме ПД в этих клетках возникает в результате прихода импульсов от более часто разряжающихся верхних отделов. Иначе говоря, собственный автоматизм нижних отделов не успевает проявиться. Так как наивысшая частота у СА, то он является ведущим пейсмекером первого порядка.
Соподчинение водителей ритма. Все отделы проводящей системы способны спонтанно генерировать импульсы, но с различной частотой. Самой большой частотой обладают клетки СА. Все другие имеют существенно меньшую частоту разрядов, причем, чем дальше отдел проводящей системы расположен от СА, тем его частота ниже. Благодаря этому в норме ПД в этих клетках возникает в результате прихода импульсов от более часто разряжающихся верхних отделов. Иначе говоря, собственный автоматизм нижних отделов не успевает проявиться. Так как наивысшая частота у СА, то он является ведущим пейсмекером первого порядка.
В том случае, если возбуждение в СА не возникает, либо, например, при синоатриальной блокаде, не может перейти на желудочки, роль пейсмекера второго порядка берет на себя АВ-узел с частотой импульсов 40-60 в мин.
Рис. 21. Лигатуры Станниуса
В случае полной поперечной блокады предсердия и желудочки начинают сокращаться независимо друг от друга – предсердия в ритме СА-узла, а желудочки с частотой 30-40 в мин – с частотой, присущей пейсмекеру третьего порядка. Это явление называют диссоциацией.
Доказать, что синоатриальный узел является местом возникновения возбуждения в сердце (пейсмекером) можно или электрофизиологически, или опытами с ограниченным охлаждением синоатриального узла. Местное охлаждение вызывает замедление или остановку сердца, тогда как действие холода на другие участки сердца неэффективно. По мускулатуре предсердий возбуждение распространяется со скоростью около 1 м/ сек, и доходит до элементов атриовентрикулярного узла. При переходе через последний отмечается задержка возбуждения, так что на пучок Гисса возбуждение передается лишь через 0,04-0,05 сек после того, как оно дошло до атриовентрикулярного узла. В течение этой атриовентрикулярной задержки систола предсердий уже заканчивается. Таким образом, систола желудочков начинается только после окончания систолы предсердий.
Автоматия сердца. Сердце, даже после того, как оно вырезано из тела, может ритмически сокращаться некоторое время, Способность тканей возбуждаться под влиянием импульсов, возникающих в них самих, принято называть автоматизмом.
Автоматию сердца проще всего наблюдать на изолированном сердце лягушки, помещенном в физиологический раствор. При перфузии коронарных артерий можно видеть автоматию и на сердце теплокровного животного. Томский ученый Кулябко в 1902 г. впервые оживил сердце ребенка через несколько часов после смерти, Андреев - сердце взрослого через 2 суток после смерти.
Автоматия разных отделов сердца неодинакова. Особенностью проводящей системы сердца является способность каждой клетки самостоятельно генерировать возбуждение. Существует так называемый градиент автоматии, выражающийся в убывающей способности к автоматии различных участков проводящей системы по мере их удаления от синусно-предсердного узла, генерирующего импульса с частотой до 60—80 в минуту (закон градиента автоматии Гаскелла).
Доказывается - лигатурами Станниуса или наблюдением за сокращением кусочков мышцы из разных отделов сердца. В нормальных физиологических условиях функционирует лишь один очаг автоматии - синоатриальный узел. Водители ритма, находящиеся в других отделах сердца, не генерируют импульсов. Их автоматия подавлена более частым ритмом синоатриального узла (способность к усвоению более частого ритма). После наложения лигатуры на предсердия наступает преавтоматическая пауза, после которой начинает проявляться автоматия атриовентрикулярного узла.
Ритмическое сокращение можно наблюдать и на трипсинизированном сердце (в культуре ткани). Отдельные мышечные клетки сокращаются и генерируют импульсы. При этом со временем клетки собираются в комочки и начинают сокращаться в ритме самого частого темпа, свойственного данным клеткам.
Электрофизиологические исследования, проведенные с помощью внутриклеточных микроэлектродов, показали, что в промежутке между двумя сокращениями в диастолу в автоматически возбуждающихся клетках происходит постепенное уменьшение мембранного потенциала (медленная диастолическая деполяризация). Когда разность потенциалов уменьшается до критической, внезапно возникает крутой сдвиг электрического заряда и генерируется ПД. Чем быстрее изменяется мембранный потенциал, тем чаще автоматический ритм. Считают, что это зависит от особенностей проницаемости поверхностной мембраны мышечного волокна, и от наличия в ней циклических биохимических процессов, ведущих к периодическому изменению проницаемости.
Электрическая активность клеток миокарда. В естественных условиях клетки миокарда находятся в состоянии ритмической активности (возбуждения), поэтому об их потенциале покоя можно говорить лишь условно. У большинства клеток он составляет около 90 мВ и определяется почти целиком концентрационным градиентом ионов К+.
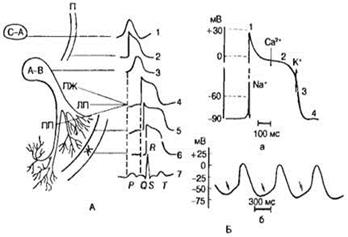 Рис. 22. Различные типы потенциалов действия сердечных клеток.
Рис. 22. Различные типы потенциалов действия сердечных клеток.
Потенциалы действия (ПД), зарегистрированные в разных отделах сердца при помощи внутриклеточных микроэлектродов, существенно различаются по форме, амплитуде и длительности (рис. 22, А). На рис. 22, Б схематически показан ПД одиночной клетки миокарда желудочка. Для возникновения этого потенциала потребовалось деполяризовать мембрану на 30 мВ. В ПД различают следующие фазы: быструю начальную деполяризацию — фаза 1; медленную реполяризацию, так называемое плато — фаза 2; быструю реполяризацию — фаза 3; фазу покоя — фаза 4. Фаза 1 в клетках миокарда предсердий, сердечных проводящих миоцитов (волокна Пуркинье) и миокарда желудочков имеет ту же природу, что и восходящая фаза ПД нервных и скелетных мышечных волокон — она обусловлена повышением натриевой проницаемости, т. е. активацией быстрых натриевых каналов клеточной мембраны. Во время пика ПД происходит изменение знака мембранного потенциала (с —90 до +30 мВ).
В клетках рабочего миокарда (предсердия, желудочки) мембранный потенциал (в интервалах между следующими друг за другом ПД) поддерживается на более или менее постоянном уровне. Однако в клетках синусно-предсердного узла, выполняющего роль водителя ритма сердца, наблюдается спонтанная диастолическая деполяризация (фаза 4), при достижении критического уровня которой (примерно —50 мВ) возникает новый ПД (см. рис. 22, Б). На этом механизме основана авторитмическая активность указанных сердечных клеток.
Биологическая активность этих клеток имеет и другие важные особенности: 1) малую крутизну подъема ПД; 2) медленную реполяризацию (фаза 2), плавно переходящую в фазу быстрой реполяризации (фаза 3), во время которой мембранный потенциал достигает уровня —60 мВ (вместо —90 мВ в рабочем миокарде), после чего вновь начинается фаза медленной диастолической деполяризации. Сходные черты имеет электрическая активность клеток предсердно-желудочкового узла, однако скорость спонтанной диастолической деполяризации у них значительно ниже, чем у клеток синусно-предсердного узла, соответственно ритм их потенциальной автоматической активности меньше.
Ионные механизмы генерации электрических потенциалов в клетках водителя ритма полностью не расшифрованы. Установлено, что в развитии медленной диастолической деполяризации и медленной восходящей фазы ПД клеток синусно-предсердного узла ведущую роль играют кальциевые каналы. Они проницаемы не только для ионов Са2+, но и для ионов Na+. Быстрые натриевые каналы не принимают участия в генерации ПД этих клеток.
Способность клеток миокарда в течение жизни человека находиться в состоянии непрерывной ритмической активности обеспечивается эффективной работой ионных насосов этих клеток. В период диастолы из клетки выводятся ионы Na+, а в клетку возвращаются ионы К+. Ионы Са2+, проникшие в цитоплазму, поглощаются эндоплазматической сетью. Ухудшение кровоснабжения миокарда (ишемия) ведет к обеднению запасов АТФ и креатинфосфата в миокардиальных клетках; работа насосов нарушается, вследствие чего уменьшается электрическая и механическая активность миокардиальных клеток.
Таким образом, наличие проводящей системы обеспечивает ряд важных физиологических особенностей сердца: 1) ритмическую генерацию импульсов (потенциалов действия); 2) необходимую последовательность (координацию) сокращений предсердий и желудочков; 3) синхронное вовлечение в процесс сокращения клеток миокарда желудочков (что увеличивает эффективность систолы).
Экстрасистолы. Сокращение (систола) миокарда продолжается около 0,3 с, что по времени примерно совпадает с рефрактерной фазой. Следовательно, в период сокращения сердце неспособно реагировать на другие раздражители. Наличие длительной рефрактерной фазы препятствует развитию непрерывного укорочения (тетануса) сердечной мышцы, что привело бы к невозможности осуществления сердцем нагнетательной функции.
Раздражение, нанесенное на миокард в период расслабления (диастолы), когда его возбудимость частично или полностью восстановлена, вызывает внеочередное сокращение сердца — экстрасистолу. Наличие или отсутствие экстрасистол, а также их характер определяется при регистрации электрокардиограммы.
При некоторых патологических состояниях сердца правильный ритм эпизодически или регулярно нарушается внеочередной систолой, которая называется экстрасистолой.
Если внеочередное возбуждение возникает (или в эксперименте мы раздражаем) в области синоатриального узла, то в случае, когда этот импульс попадает в период после окончания рефрактерности синусового возбуждения, наступает внеочередное возбуждение синусного узла и внеочередное сокращение всего сердца (синусовая или предсердная экстрасистола). Пауза, следующая за такой экстрасистолой, длится столько же времени, сколько обычная пауза, так как после внеочередного возбуждения синусный узел автоматически генерирует очередной импульс с обычным ритмом.
Если же внеочередное возбуждение возникает в водителях ритма правого или левого желудочков, или в атриовентрикулярном узле, то такое возбуждение не отражается на автоматии синоатриального узла, и этот узел своевременно посылает свой очередной импульс, который достигает желудочков, когда они еще находятся в рефрактерном состоянии после экстрасистолы. Поэтому миокард не отвечает на очередной импульс из синусного узла, и ждет следующего больший, чем обычно, период времени. Возникает т.н. компенсаторная пауза, наличие которой является отличительной особенностью атриовентрикулярной (желудочковой) экстрасистолы.
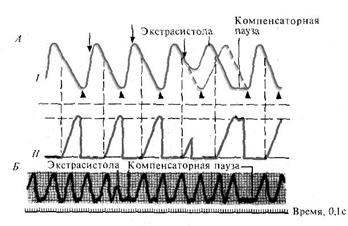 Рис. 23. Предсердная (I) и атриовентрикулярная (II) экстрасистолы.
Рис. 23. Предсердная (I) и атриовентрикулярная (II) экстрасистолы.
При экстрасистолах, возникающих в правом или левом желудочках, форма их на ЭКГ разная и не похожа на нормальный желудочковый комплекс. Если же экстрасистола возникает из атриовентрикулярного узла, то форма QRST остается обычной. Однако в этом случае меняется форма зубца Р. Он направлен вниз, так как возбуждение из атриовентрикулярного узла на предсердия распространяется снизу. Кроме того, в зависимости от того, в какой части узла возникает возбуждение, зубец Р может или предшествовать, или сливаться, или быть после желудочкового комплекса. Источником экстрасистолии могут быть патологические очаги в различных отделах сердца, или импульсы из нервной системы.
Дата: 2019-04-23, просмотров: 556.