Мир живых организмов на Земле чрезвычайно разнообразен. В настоящее время на нашей планете насчитывается несколько миллионов животных, растений грибов и микроорганизмов.
Каждый вид можно описать совокупностью характерных черт, свойств, которые называются признаками вида. Признаки, благодаря которым можно отличить один вид от другого называются критериями вида. Наиболее часто пользуются шесть критериев вида: морфологический, физиологический, географический, экологический, генетический и биохимический.
Морфологический критерий, основанный на внешнем и внутреннем сходстве особей, самый простой, однако он не срабатывает при различении видов, имеющих морфологическое сходство.
Физиологический критерий заключается в сходстве жизненных процессов, в первую очередь, возможности скрещивания между особями одного вида и образования плодовитого потомства. Между разными видами существует физиологическая изоляция, но вместе с тем между некоторыми видами скрещивание возможно и, при этом могут образовываться плодовитые гибриды.
Географический критерий основан на том, что каждый вид занимает определенную территорию или акваторию, характеризуется определенным ареалом. Многие виды занимают отдельные ареалы, но огромное количество видов имеют накладывающиеся ареалы.
Экологический критерий основан на том, что каждый вид может существовать только в определенных абиотических и биотических условиях (занимает свою экологическую нишу). Вместе с тем существуют виды, не имеющие строгой экологической привязанности. Это синантропные виды (обитающие рядом с человеком) – тараканы, мухи, мыши, крысы, домашние животные, комнатные растения.
Генетический критерий обусловлен различием видов по кариотипам. Для большинства видов характерен строго определенный набор хромосом. Однако в пределах одного вида могут встречаться особи с разным числом хромосом (диплоидным, тетраплоидным).
Биохимический критерий объединяет вид по совокупности биохимических структур и реакций. Но применение этого критерия трудоемко и существует внутривидовая изменчивость практически всех показателей.
Каждый вид занимает различные ареалы. Особи одного вида распространены в ареале неравномерно, что обусловлено различиями условий жизни, сложившихся в различных его участках (климатические особенности, условия питания и т.д.). Особи любого вида живут не по одиночке, а группами, внутри которой сформировались сложные взаимоотношения.
В природе представители отдельных видов, как правило, объединены в определенные совокупности – популяции.
Популяцией называют группу особей одного вида, обладающих способностью свободно скрещиваться и неограниченно долго поддерживать свое существование в определенном местообитании. Популяция - это некоторое единство, которое определяется общностью занимаемой особями территории (или акватории), а также общностью их происхождения, сходством строения и поведения. Например: все особи одного вида, обитающие в небольшом озере, или все деревья одного вида в лесу.
Популяция представляет один из уровней организации жизни в биосфере и ее составляют особи одного вида, которые проживают на общей территории. Различают экологическое и генетическое определение популяции.
Популяция в экологическом плане это все особи (животных или растений), которые входят в состав одного вида на какой-то изолированной части природного обитания. В геоботанике для обозначения популяции растений, которые образуют различные фитогеоценозы, употребляют термин «ценопопуляция».
Генетики определяют популяцию как группу особей, которые скрещиваются между собой и имеют единую генетическую основу. Каждая популяция выполняет четко определенную функцию в том биогеоценозе, в котором она распространена. На состояние популяции влияют процессы, которые происходят в самой популяции, так и соседних популяциях. Все популяции подразделяются на определенные категории по способности к самоподдержанию.
Наиболее близким по значению к значению термина «популяция» является понятие «племя». Следовательно, популяции состоят из одинаковых организмов, совместно населяющих определенные участки и связанных между собой различными взаимоотношениями, которые обеспечивают им устойчивое существование в данной природной среде.
Слово «популяция» происходит от латинского «популюс» - народ, население. Экологическую популяцию, таким образом, можно определить как население одного вида на определенной территории.
Члены одной популяции оказывают друг на друга не меньшее взаимодействие, чем абиотические факторы среды или другие обитающие совместно виды организмов. В популяциях проявляются в той или иной степени все формы связей, характерные для межвидовых отношений, но наиболее ярко выражены мутуалистические (взаимно полезные) и конкурентные. Во всех случаях в популяциях действуют законы, позволяющие таким образом использовать ограниченные ресурсы среды, чтобы обеспечить оставление потомства. Достигается это в основном через количественное изменение населения. Популяции многих видов обладают свойствами, позволяющими им регулировать свою численность.
Смешиванию популяций препятствуют различные барьеры: абиотические – горы, реки, моря, характер почвы и т.д.; биотические – хронологические (из-за расхождения в жизненных циклах, чаще в период размножения); этологические – характерное поведение животных, проявляющееся в охране территории от чужих особей; биогеоценотические, проявляющиеся в преимуществе особей популяции в местах расселения перед эмигрантами; антропогенные – различные уровни загрязнения воздуха, воды, почвы и другие антропогенные воздействия, вызывающие различные адаптации к ним живых организмов.
Каждая популяция выполняет строго определенную функцию в биологическом сообществе. На состояние популяции влияют процессы, которые происходят как внутри ее, так и соседних популяциях. Можно выделить следующие разновидности популяций:
1. Независимые популяции имеют достаточно высокий потенциал размножения, благодаря которому постоянно восполняются потери особей без поступления извне.
2. Полунезависимые популяции. Они могут существовать при размножении собственных особей, но поступление особей извне значительно улучшает состояние данного сообщества.
3. Зависимые популяции могут существовать только при условии поступлении особей из соседних популяций, так как рождаемость в них не компенсирует потерь.
4. Псевдопопуляции распространены в той местности, где группа особей не имеет возможности размножаться на данной территории. Такие популяции возникают в результате миграции особей из соседних популяций. Это популяции, которые возникают за пределами распространения или в биотопах, которые еще не полностью заселены. Появление их связано с периодическим возникновением благоприятных условий. При ухудшении условий среды обитания такие популяции исчезают
5. Гемипопуляции - состояние сообществ, когда дифференцируются потребности на разных фазах жизненного цикла определенного вида. При этом популяция состоит из дифференцированных компонентов, приспособленных к определенным условиям среды, пространства, которые циклично идут один за другим (фазы развития комара и других насекомых происходят в различных условиях).
Каждая популяция характеризуется определенными признаками: ареалом, численностью и плотностью, возрастной и половой структурой.
Любая популяция занимает определенную территорию или акваторию, которая называется популяционным ареалом, который может быть различной величины. Размеры ареала в значительной мере зависят от степени подвижности особей: чем меньше подвижность особей, тем меньше величина популяционного ареала.
Одной из структурных характеристик популяций является распределение особей в популяционном ареале. Выделяют три типа распределения особей в популяции: а) случайное; б) равномерное; в) групповое. Случайное распределение особей наблюдается при однородных условиях среды и когда особи не объединены в группы. Равномерное – при антагонистических отношениях между особями и присутствии конкуренции. Групповое характерно для популяций, где биологические особенности обуславливают создание групп, скоплений. Скопление особей в определенных местах ареала может возникать в связи с процессами размножения, под влиянием суточной и сезонной динамики факторов окружающей среды, вследствие изменения местных условий, отмечается у животных с развитыми элементами сознания, которые определяют поведение.
Поддержание оптимальной в данных условиях численности вида называют гомеостазом популяции. Гомеостатические возможности популяций по-разному выражены у различных видов. Осуществляются они через взаимодействия особей.
Таким образом, популяции, как групповые объединения, обладают рядом специфических свойств, которые не присущи каждой отдельной особи. Групповые особенности - это основные характеристики популяций. К ним относятся:
1) численность - общее количество особей на выделяемой территории;
2) плотность - среднее число особей на единицу площади или объема, занимаемого популяцией пространства; плотность популяции можно выражать также через массу членов популяции в единице пространства;
3) рождаемость - число новых особей, проявившихся за единицу времени в результате размножения;
4) смертность - показатель, отражающий количество погибших в популяции особей за определенный отрезок времени;
5) прирост популяции - разница между рождаемостью и смертностью; прирост может быть как положительным, так и отрицательным;
6) темп роста - средний прирост за единицу времени.
Каждая популяция характеризуется определенной численностью. У насекомых и растений открытых пространств численность в популяциях может достигать тысяч и миллионов особей. В то же время в популяции некоторых млекопитающих может насчитываться несколько сотен или тысяч особей.
Численность популяций может сильно варьировать, однако она не может быть ниже некоторых пределов. Сокращение численности за эти пределы может привести к вымиранию популяции. Полагают, что при численности менее нескольких сотен особей какие-либо случайные причины (пожары, наводнения, обильные снегопады, сильные морозы и др.) могут сократить численность популяции настолько, что оставшиеся особи не смогут встретиться и оставить потомство. Рождаемость перестанет покрывать естественную убыль и оставшееся особи в течение короткого промежутка времени вымрут.
Количество особей на единицу площади - плотность популяции находится в тесной связи с величиной ареала и численностью. Хищники имеют более низкий показатель плотности, чем травоядные виды. Плотность популяции является максимальной, когда увеличение ее численности на данной территории не может поддерживаться экосистемой. Минимальная плотность определяется наименьшей численностью особей на единицу площади, при которой размножение не представляется возможным.
Возрастная структура популяции – соотношение групп особей разного возраста, является одной из важнейших характеристик популяции. Этот показатель характеризует способность сообщества к размножению в данный момент. В возрастной структуре популяции важное место занимают такие признаки как рождаемость и смертность.
Рождаемость – это способность популяции к поддержанию и увеличению численности. Она характеризует появление новых особей в результате рождения любого организма, независимо от того, как это происходит (они рождаются, вылупливаются из яиц, вырастают из семян или появляются в результате деления).
Максимальная рождаемость (абсолютная) – теоретический максимум скорости образования новых особей в идеальных условиях. Максимальная рождаемость постоянна для данной популяции.
Реальная рождаемость (или экологическая) определяет увеличение численности популяции при фактических условиях окружающей среды. Эта величина непостоянна и колеблется в зависимости от размеров и возрастного состава популяции и условий окружающей среды. Преимущественно рождаемость выражают как скорость, которая определяется числом рожденных особей за единицу времени на одну особь популяции (удельная рождаемость).
Смертность отражает гибель особей в популяции. Аналогично рождаемости, смертность можно выразить количеством особей, которые погибли за определенный период (количество смертей за единицу времени). Она имеет закономерный характер и неодинакова для особей различного пола и возраста; причины ее и интенсивность закономерно изменяются при смене среды и самой популяции. Степень смертности определяет продолжительность жизни и последовательность отмирания определенных генераций, а так же возрастной состав популяции.
От величины гибели особей в разных фазах развития зависит характер смертности популяции. Он может быть представлен различными типами кривых (Рис. 3)
Кривая 1 типа встречается тогда, когда на протяжении всей жизни смертность ничтожно мала, резко возрастая в конце нее. Данный тип характерен для насекомых, которые обычно гибнут после кладки яиц (называют кривой «дрозофилы»), к ней приближаются кривые выживаемости человека в развитых странах, а так же некоторых крупных млекопитающих.
Кривая 3 типа – это случаи массовой гибели особей в начальный период жизни. Гидробионты и некоторые другие организмы, не проявляющие заботы о потомстве, выживают за счет огромного числа личинок, икринок, семян и т.п.
Численность
 |


 1
1
2
3
 Время
Время
Рис. 3. Различные типы кривых выживания (объяснения в тексте).
Кривая 2 типа (диагональная) характерна для видов, у которых смертность остается примерно постоянной в течение всей жизни. Такое распределение смертности не столь уж редкое явление среди организмов. Встречаются они среди рыб, пресмыкающихся, птиц, многолетних травянистых растений.
Реальные кривые выживания чаще всего представляют собой некоторую комбинацию указанных выше «основных типов». Например, у крупных млекопитающих (и у людей живущих в развивающихся странах) кривая 1 типа вначале круто падает за счет повышенной смертности сразу после рождения.
К важнейшим характеристикам популяций относится так же половое соотношение особей в сообществе. Соотношение полов – это отношение количества самцов к количеству самок или количество самцов к общей численности популяции. Это играет роль только у раздельнополых организмов. Большинство высших растений и некоторые низшие животные являются гермафродитами и для них эти проблемы не существуют. Высшие животные (позвоночные и часть беспозвоночных) размножаются половым способом, в котором берут участие двое родителей. Так как млекопитающие могут оплодотворять нескольких самок, для эволюции их популяции большее значение имеет численность взрослых самок. Только отдельные виды образуют на период размножения отдельные пары, которые могут сохраняться до конца жизни одного из партнеров. Соотношение полов у высших животных приобретает практическое значение (куры, коровы и другие домашние животные).
Популяции свойственна определенная организация. Распределение особей по территории, соотношения групп по полу, возрасту, морфологическим, физиологическим, поведенческим и генетическим особенностям отражают структуру популяции. Она формируется, с одной стороны, на основе общих биологических свойств вида, а с другой - под влиянием абиотических факторов среды и популяций других видов. Структура популяций имеет, следовательно, приспособительный характер. Разные популяции одного вида обладают как сходными особенностями структуры, так и отличительными, характеризующими специфику экологических условий в местах их обитания.
Таким образом, кроме адаптивных возможностей отдельных особей, население вида определенной территории характеризуется еще и приспособительными чертами групповой организации, которые являются свойствами популяции как надиндивидуальной системы. Адаптивные возможности вида в целом как системы популяций значительно шире приспособительных особенностей каждой конкретной особи.
Экология популяций
Для понимания роли вида в биогеоценозе (экосистеме) необходимо знать особенности не отдельных особей вида, а больших групп, которые взаимодействуют между собой и окружающей средой. Состав популяции определяется не случайными комбинациями индивидуумов, а определенными взаимосвязями вида с комплексом условий окружающей среды в процессе поддержания видовой численности, а поэтому каждая популяция кроме общих видовых особенностей имеет специфические.
Структура популяций.
Численность и плотность популяции.
Количество особей, которые составляют популяцию (численность), а также плотность особей на единицу площади – это формальные элементы структуры популяции. Основными параметрами популяции являются плотность, возрастное и генетическое разнообразие (генофонд).
У редких или исчезающих видов контролируется их численность. В первую очередь это касается видов, которые охраняются. Полный учет видов растений или животных осуществляется периодически и является основой для оценки состояния этих видов. Так проводится инвентаризация крупных растений (тисов) и животных (лоси, тигры, бизоны и др.). Собранная информация о динамике численности используется для разработки мероприятий по их охране. Для определения численности и плотности популяции используются много разнообразных методов: тотальный учет, метод пробных участков, метод отлова, метод метки и последующего отлова, метод взятия проб (используется для неподвижных организмов, например деревьев).
Пространственная структура популяции.
Популяции изменяются во времени и пространстве. Известно, что особи никогда не размещаются на территории расселения равномерно. Выделяю 3 типа пространственной структуры популяции:
- случайные;
- равномерные;
- групповые неравномерные.
Динамика популяций.
Динамику численности популяции и механизмы ее поддержания важно понимать для того, чтобы можно было регулировать развитие сообществ, особенно полезных и вредителей для человека. Еще в 18 веке было установлено, что численность популяции растет по закону геометрической прогрессии, а в конце 18 века Томас Мальтус (1766-1834) выдвинул свою известную теорию роста народонаселения в геометрической прогрессии. Эта закономерность отражена на
рис. 4.
 | | ||||
 | |||||
 N lnN N
N lnN N
 | |||
 | |||

 t t t
t t t
а) б) в)
Рис. 4. Экспоненциальный рост гипотетической популяции одноклеточного организма, делящегося каждые 4 часа (а, б) и логистическая модель роста популяции (в).
Экспоненциальный рост возможен только тогда, когда условия размножения остаются неизменными. Такая ситуация невозможна в природе. Если бы это было не так, то, например, обычные бактерии могли бы дать такую массу органического вещества, которая могла покрыть весь земной шар слоем толщиной в два метра за несколько суток.
Воздействие экологических факторов на скорость роста популяции может довести ее численность до стабильной, либо уменьшить, т.е. экспоненциальный рост замедляется либо останавливается. В природе так и происходит: экспоненциальный рост наблюдается какое-то достаточно короткое время, после чего ограничивающие факторы его стабилизируют и дальнейшее развитие популяции идет по логистической модели (Рис. 4в)
Выделяют несколько фаз численности популяций. В первой фазе отмечается общий прирост численности сообщества, а во второй – уменьшается численность. В фазе равновесия численность популяции остается практически неизменной. Однако такое состояние относительно редкое явление. Наиболее часто в природе численность популяции колеблется относительно какого-то определенного среднего значения. Динамика численности популяции зависит от внутренних и внешних факторов. К внешним факторам относятся факторы окружающей среды и биоценотические взаимоотношения. Различают следующие типы динамических изменений численности популяций:
- демонстративный тип характеризуется периодом интенсивного увеличения численности в геометрической прогрессии. После достижения максимума количество особей уменьшается. Продолжительность периода полного цикла таких изменений зависит от биологических особенностей вида;
- гиперболический тип присущ популяциям, которые осваивают местность с ограниченными пищевыми ресурсами. Популяция увеличивается при освоении пространства и ресурсов среды, после чего она переходит в состояние равновесия.
- циклический тип характеризует популяцию, которая находится в неуравновешенных, несбалансированных условиях среды;
- стабильный тип динамики численности популяции присущ тем сообществам, которые находятся в установившихся условиях и имеют совершенные регуляторные механизмы. Такой тип распространен в природе редко.
Гомеостаз популяций
Устойчивость популяции, ее относительная самостоятельность и «индивидуальность» зависят от того, насколько структура и внутренние свойства популяции позволяют приспособиться к изменяющимся условиям окружающей среды. Именно в поддержании динамического равновесия со средой и заключается принцип гомеостаза популяции как целостной биологической системы. При всем многообразии проявления механизмов популяционного гомеостаза их можно сгруппировать в три важнейших группы: 1 – поддержание адаптивного характера пространственной структуры, 2 – поддержание генетической структуры, 3 - регуляция плотности.
Пространственная структура популяции.
Видовые особенности питания, размножения, отношения к абиотическим факторам формируют, свойственный виду, характер использования территории и тип отношений внутри сообщества (социальных отношений). Это определяет видовой тип пространственно-этологической структуры популяции. Наиболее важные показатели этого является общий характер использования территории, привязанность к определенной местности, особенности распределения особей в пространстве. Видовой тип пространственной структуры включает закономерные, периодические повторяющиеся (например, сезонные) изменения перечисленных параметров.
Среди механизмов поддержания пространственной структуры популяции можно выделить несколько определяющих:
- механизмы «индивидуализации» территории;
- механизмы поддержания иерархии.
Преимущества обитания на постоянном участке могут быть реализованы только при условии использования ее постоянно обитающими на ней особями или группами. Особи-резиденты привязаны к территории участка системой знакомых ориентиров, выход за пределы которой вызывает ориентировочную реакцию, служащую стимулом возвращения на участок. Но такое поведение не гарантирует от проникновения на территорию резидентов других особей того же вида. Это определяется комплексом поведенческих реакций, свойственным оседлым животным с интенсивным использованием территории: территориальная агрессия, маркирование территории, регуляция территориального поведения.
Территориальная агрессия в той или иной форме свойственна всем видам, имеющим участки обитания. Биологически важно, что при территориальном конфликте победителем в большинстве случаев оказываются особи, на участке которых произошла встреча (особь-резидент, «хозяин» данного участка).
Маркирование территории представляет собой не связанный с агрессией способ индивидуализации участка. Способы маркирования участка разнообразны. У тропических рыбок яркая окраска с броским рисунком делает присутствие рыбы достаточно ощутимым сигналом занятости территории. У птиц отчетливо выражена акустическая маркировка участков в виде песни и других звуковых сигналов. У большинства млекопитающих наиболее важное значение в маркировании территории имеют запаховые метки. Носителем запаха служит моча, экскременты, но наиболее специализированное значение имеют секреты особых желез, свойственных многим территориальным видам.
Регуляция территориального поведения, маркировка и территориальная агрессия связана с физиологией животных, в первую очередь с состоянием генеративной системы и соответствующим гормональным фоном в организме. Функционально активное состояние половой системы – мощный стимул в проявлении всего комплекса территориального поведения. У многих видов рептилий защита и маркировка территории наблюдается только в период размножения и интенсивность проявления территориального поведения зависит от стадии репродуктивного цикла. Такой механизм регуляции территориального поведения биологически оправдан: именно в период размножения животные наиболее привязаны к территории и индивидуализация ее особенно важна.
Механизмы поддержания иерархии.
Формирование и поддержание иерархической структуры взаимоотношений – важный процесс, определяющий пространственную целостность и функциональную стабильность внутрипопуляционных групп и популяции в целом. В основе функциональной интеграции особей в популяциях лежит система соподчинения животных, основанная на разнокачественности особей в составе популяции.
В основе формирования иерархии лежат сложные комплексы поведения и физиологические реакции. Среди последних, у млекопитающих важное значение имеет неспецифическая адаптивная реакция, возникающая в ответ на действие угнетающих (стрессирующих) факторов, получившая название «реакции напряжения» или стресса.
В многочисленных опытах с грызунами установлено, что начало формирования групп из раннее незнакомых особей у большинства видов сопровождается вспышкой агрессии. В процессе возникающих схваток и агрессивного демонстративного поведения, часть особей через некоторое время выбывает из борьбы; эти особи занимают соответствующе подчиненные ранги в иерархии.
Установлено, что становление системы доминирования имеет значение также такие фундаментальные свойства, как врожденный уровень агрессивности. В опытах с домовыми мышами, курами и некоторыми другими животными выявлена корреляция индивидуальной агрессивности с типологическими особенностями ЦНС: агрессивность выше у животных с сильными уравновешенными и подвижными нервными процессами.
Имеется большое число наблюдений и экспериментов, показывающих роль индивидуального опыта в возможности занять высокоранговое положение. Так роль «бойцовского» опыта в исходе агрессивных схваток описана у некоторых рыб. В опытах с белыми мышами установлено, что самцы, натренированные на победу в схватках, могли не только проникать в состав сложившихся групп, несмотря на встречную агрессию, но и занимать в них доминирующее положение.
У стадных животных физиологические основы выделения вожака, вероятно, близки к механизмам становления доминантов у оседлых видов. Хотя вожак-доминант выделяется в большинстве случаев в процессе драк и угроз, физическая сила – не главное, что определяет ранг животного. По-видимому у стадных животных определенную роль в становлении вожака и устойчивом осуществлении им своих функций играют генетически детерминированные индивидуальные особенности физиологии и высшей нервной деятельности.
Взрослые особи, имеющие значительный индивидуальный опыт, в том числе и опыт побед в схватках, формируют особый стереотип поведения, дающий преимущества как им самим в борьбе с менее опытными, так и стае в целом в ситуациях требующих быстрой приспособительной реакции.
В сложившихся стадах с вожаками поддержание структуры и организации деятельности группы осуществляется на основе стереотипных форм поведения, ориентированных на высокоранговых особей, как путем подражания, так и с помощью сложной системы реакции на коммуникативные сигналы с их стороны.
Логистическая модель роста популяции предполагает наличие некой равновесной численности и плотности. В этом случае рождаемость и смертность должны быть равны, т.е. должны действовать факторы, изменяющие либо рождаемость, либо смертность.
Факторы, регулирующие плотность популяции, делятся на зависимые и независимые от плотности. Зависимые изменяются с изменением плотности, а независимые остаются постоянными при ее изменении. Практически, первые – это биотические, а вторые – абиотические факторы.
Поддержание генетической структуры
Генетическое многообразие в популяции обеспечивает преимущества в приспособлении сообщества к изменяющимся условиям окружающей среды. Если весь диапазон индивидуальной изменчивости адаптивных свойств в популяции представить в виде вариационной кривой, то ее средние характеристики отразят «настройку» этих свойств на средние, наиболее типичные и устойчивые условия среды. При изменении этих условий более приспособленными, адаптированными оказываются особи, не входящие в среднюю группу, а отклоняющиеся от нее в сторону, адекватную измененным условиям. Именно эти особи и обеспечивают выживание и последующее восстановление популяции. В дальнейшем, если возникшие условия являются стабильными, в популяции вступает в силу отбор особей приспособленных к условиям окружающей среды. Это экологический процесс, обеспечивающий устойчивость популяционной системы в колеблющихся условиях среды. В самом общем виде этот процесс может быть представлен как система специфических внутрипопуляционных отношений, направленных на снижение уровня генетической однородности (имбридинга) в размножающейся популяции.
В структурированных по взаимоотношениям группах животных доминирующие особи имеют преимущества в спаривании. Это особенно характерно для видов с жесткой иерархической структурой. У животных с менее жесткой системой доминирования в размножении участвуют соответственно большая доля самцов в популяции. Но элементы сексуального доминирования имеют место и в этом случае. Так, у территориальных животных из размножения исключаются особи, не сумевшие захватить и удержать участок обитания. Среди обладающих территорией самцов участие в размножении также неодинаково, что в первую очередь связано с их положением в этологической структуре внутрипопуляционных групп.
Физиологические механизмы сексуального доминирования прямо связаны с эколого-физиологическими процессами установления и поддержания иерархии. Доминанты, несомненно, представляют собой особей, наиболее успешно приспособившихся к конкретным условиям, способным четко и адаптивно реагировать на их изменения. Таким образом, сексуальное доминирование полезно на уровне популяции, как механизм закрепления полезных генотипов. Но с другой стороны, направленное уменьшение случайных скрещиваний способствует снижению генетического разнообразия (гетерозиготности) популяции. Устранение данного противоречия – задача большой биологической важности. Отмечено, что даже в условиях самой жесткой иерархии сексуальное доминирование регулярно нарушается и в размножение включается дополнительные контингенты самцов.
Закономерное нарушение доминантности при скрещивании в природе встречается очень часто. Описаны случаи, когда сексуальное доминирование нарушается типом половой избирательности самок, которые отдают предпочтение низкоранговым особям.
Доминантная структура не остается неизменной даже при самых жестких формах иерархии. Изучение различных видов млекопитающих показывают, что в сложившихся внутрипопуляционных группах доминанты регулярно сменяются. Доминирующую особь может вытеснить внедрившееся в группу новое, более сильное и агрессивное животное. В других случаях место доминанта может занять особь и той же группы в результате перераспределения рангов.
Подвижность и расселение играет у некоторых видов важную роль в поддержании генетического разнообразия. Расселение особей за пределы мест рождения известно для многих видов животных; так же широко известны различные способы распространения семян растений на расстояния, превышающие возможность опыления. Процесс этот может быть пассивным (использование течений, ветра), но может быть активным; последнее характерно для многих видов позвоночных животных.
Расселение молодняка у животных выполняет много функций и жизни популяции: захват пространства, повышение вероятности переживания неблагоприятных сезонов, смягчение территориальных отношений и т.д. Наряду с этим в результате рассеивания в пространстве происходит регулярное «перемешивание» особей в популяции, что облегчает панмиксию (свободное скрещивание между особями) и ведет к обогащению генофонда популяции через снижение инбридинга (близкородственного скрещивания).
У молодых животных стимул к миграции, по-видимому, закодирован в генетической программе онтогенеза, и он часто усиливается внутрисемейной агрессией, возникающей к моменту возмужания молодняка.
Мощным фактором, нарушающим случайный характер спаривания у высших животных, является избирательность по отношению к половым партнерам. Половая избирательность может быть выражена в разных формах и неодинаково представлена у самцов и самок, но при всем многообразии форм выполняет три важные биологические функции:
- предотвращает слишком отдаленные (между видами и подвидами) скрещивания;
- закрепляет адаптивные свойства в пределах данной популяции;
- снижает частоту близкородственных скрещиваний.
Две последние из этих функций имеют важное экологическое значение: избирательное отношение к потенциальным половым партнерам способствует устойчивости популяции через поддержание оптимального типа ее генетической структуры. У млекопитающих механизмы избирательности основываются на способности к запаховой дискриминации особей.
В поддержании генетического разнообразия популяции играет важное значение спаривание особей, принадлежащих разным возрастным группам (возрастной кросс). У многих млекопитающих самцы и самки либо достигают половой зрелости в разном возрасте, либо в силу особенностей социальных взаимоотношений в сообществе самцы (а иногда и самки) не сразу реализуют свои возможности участия в размножении. Разновозрастные пары широко распространены. Структурированность популяций, определяя сексуальное преимущество высоко ранговых самцов, имеет и иное значение: производителями становятся в первую очередь самцы с полезными для популяции свойствами. Однако спариваются они с самками разных возрастов, что обеспечивает большое разнообразие популяционного генофонда и поддержание высокой степени гетерозиготности особей в потомстве.
Важными механизмами гомеостаза популяции является регуляция плодовитости и плотности. В наиболее простом виде плотность особей в популяции определяется обеспеченностью пищей. Однако непосредственное регулирование численности количеством пищи встречается в природе не очень часто. При одинаковом количестве пищи, размещенной в разных объемах среды, ведущую роль играет «вместимость среды», воспринимаемой через изменение частоты контактов (прямых и информационных) между особями. Именно этот параметр через изменение плодовитости и смертности определяет уровень численности и плотности особей в популяции. Зависимость от плотности динамики плодовитости и смертности описана у представителей самых разнообразных организмов. Показано, что цианобактерии способны к синтезу внеклеточных метаболитов, которые могут влиять на рост и скорость деления клеток в культуре. Наиболее изучены эти процессы у птиц и млекопитающих. Многолетние наблюдения за популяцией большой синицы в Англии показали, что число кладки яиц и процент вылупившихся птенцов находится в обратной зависимости численности птиц.
Механизмы регуляции плотности и численности в популяции разнообразны. Большое распространение у животных имеет химическая регуляция. Особенно эта форма регулирования представлена у низших таксонов животных (а также у микроорганизмов), не обладающих иными формами коммуникаций.
Регуляция плотности и численности через поведение свойственна высшим животным. Роль поведения в популяционных процессах широко известна у многих животных. При увеличении плотности частота прямых контактов особей приводит к каннибализму. Интенсивность прореживания популяции в этом случае выступает как прямая функция плотности особей, приводя ее в соответствие с экологическими требованиями.
Регуляция через структуру. Вследствие структурированности взаимоотношений в группах часть особей испытывает состояние стресса. При повышении частоты контактов в популяциях с высокой плотностью общий уровень стресса возрастает, что приводит к снижению плодовитости. (Рис. 5)
Влияние стресса на изменение плотности популяции неоднозначно. Резкое повышение количества стрессовых гормонов может стать причиной гибели животного. Явления подобного рода регистрируются в лабораторных условиях и в природе. Такое влияние стресса на смертность в популяции реализуется относительно редко. Гораздо более вероятно увеличение смертности в силу повышения чувствительности стрессированных животных к влиянию неблагоприятных факторов. Большую роль в стимуляции стрессового состояния играет агрессивное поведение. Соответствие уровня агрессии плотности популяции показано во многих экспериментальных и полевых исследованиях. В некоторых случаях агрессия может выступать как прямой фактор ограничения численности.
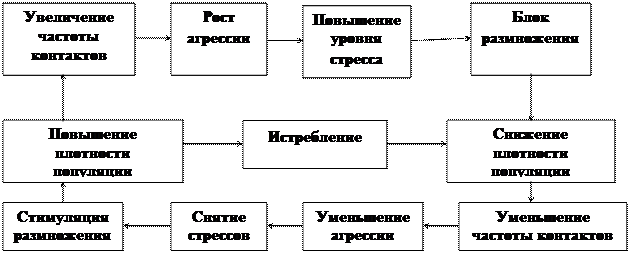 |
Рис.5. Схема популяционной авторегуляции плотности у грызунов.
Анализ динамики численности популяций различных видов показывает наличие цикличных изменений, обусловленных не только действием факторов окружающей среды, но связаны с особенностями биологии вида, его физиологией, местом в естественных экосистемах. Выделяют три основных типа динамики населения (Рис. 6).
Стабильный тип характеризуется малой амплитудой и длительным периодом численности; внешне она воспринимается как практически стабильная. Такой тип свойственен крупным животным с большой продолжительностью жизни.
Лабильный тип динамики отличается закономерными колебаниями численности с периодами порядка 5-11 лет и более значительной амплитудой (численность меняется в десятки раз). Характерны сезонные периоды размножения. Такой тип динамики характерен для животных разного вида, более мелких по размеру и с коротким периодом жизни (10-15 лет) и соответственно более ранним половым созреванием и более высокой плодовитостью, чем у представителей первого типа. К этому типу относятся грызуны, зайцеобразные, некоторые хищные.
Численность
популяции

Годы
Рис. 6 Типы динамики населения (1 - стабильный, 2- лабильный, 3 – эфемерный)
Эфемерный тип динамики отличается резко неустойчивой численностью, «вспышками размножения», при которых численность возрастает в сотни раз. Перепады от минимума к максимуму осуществляются очень быстро (иногда в течении одного сезона). Общая длина цикла обычно составляет до 4-5 лет, в течение которых пик численности занимает чаще всего не более одного года. Эфемерный тип динамики характерен для короткоживущих (не более 3 лет) видов с несовершенными механизмами индивидуальной адаптации и соответственно с высокой нормой гибели. Наиболее характерен такой тип динамики для мелких грызунов и многих видов насекомых с коротким циклом развития.
Функциональная структура популяций растений представлена системой взаимодействий, связанных с обменом генетическим материалом и процессами круговорота веществ и энергии. Осуществление этих функций в значительной степени зависит от плотности популяции. Под влиянием плотности меняется распределение питательных веществ, идущих на образование различных органов. Наиболее общее выражение этой зависимости – замедление общего роста растений при повышении плотности и отрицательное влияние плотности на развитие генеративных органов и семенную продукцию. Широко известный путь влияния плотности на популяционные процессы у растений – это явление самоизреживания, связанное с зависимой от плотности величиной смертности в ценопопуляциях.
Суммарная система регуляторных взаимоотношений различна у конкретных видов и может выражаться зависимой от плотности смертностью, а также различной реакцией отдельных элементов популяции на действие внешних факторов. В результате регуляторных процессов открывается возможность компенсировать уменьшение плотности увеличением индивидуального роста растений. В результате популяция в целом способна в значительном диапазоне изменения плотности сохранять относительно устойчивую величину биомассы. Эта закономерность («константность конечного урожая») установлена для древесных пород и для большого числа однолетних видов растений.
Вопросы для самоконтроля:
1. Характеристика биологического вида, его критерии.
2. Понятие популяции, их разновидности.
3. Групповые характеристики популяций, типы кривых выживания.
4. Структура популяции (численность, плотность, пространственное распределение).
5. Динамика популяций.
6. Гомеостаз популяций (пространственной структуры, иерархии, генетической структуры).
7. Механизмы регуляции плодовитости и плотности в популяциях.
8. Динамика численности популяций.
Биоценология (синэкология)
Биоценоз представляет собой эволюционно сложившуюся форму организации живых организмов, совокупность растений, животных, микроорганизмов, населяющих участок суши или водоема и характеризующиеся определенными отношениями и связями, как между собой, так и с абиотическими факторами среды. Как и популяция, сообщество имеет собственные свойства, проявляющиеся только при изучении его самого, как, например, видовое разнообразие, структура пищевой сети, биомасса, продуктивность (Рис. 7). Одна из главных задач экологии - выяснить взаимосвязи между свойствами и структурой (составом) сообщества, которые проявляются независимо от того, какие виды входят в него.
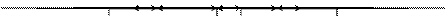
|
|

Рис. 7. Схема строения биогеоценоза (биотоп и биоценоз).
Приспособленность членов биоценоза к совместной жизни обусловлена сходством в потребности к важнейшим абиотическим условиям (свет, климатические факторы, состав почвы) и закономерным отношением друг к другу. Составными частями биогеоценоза являются фитоценоз (устойчивое сообщество растительных организмов), зооценоз (совокупность взаимосвязанных видов животных), микоценоз (сообщество грибов) и микробоценоз (сообщество микроорганизмов).
Каждый организм живет в окружении множества других организмов, вступает с ними в самые разнообразные отношения, как с отрицательными, так и с положительными для себя последствиями и в конечном счете не может существовать без этого живого окружения. Связь с другими организмами - необходимое условие питания и размножения, возможность защиты, смягчения неблагоприятных условий среды, а с другой стороны - это опасность ущерба и часто даже непосредственная угроза существования индивидуума. Всю сумму воздействий, которую оказывают друг на друга живые существа, объединяют под названием биотические факторы среды.
Биоценозы, как надорганизменный уровень организации жизни, имеет свои, отличительные от организмов, особенности:
1) Сообщества всегда возникают, складываются из готовых частей, имеющихся в окружающей среде, в отличие от организмов, которые формируются путем постепенной дифференцировки зародышей или других зачатков;
2) Части сообщества заменяемы: один вид может занять место другого со сходными экологическими требованиями без ущерба для всей системы, а части (органы и ткани) организма уникальны;
3) В организме поддерживается постоянная координация, согласованность деятельности его клеток, тканей, органов, а надорганизменная система в основном существует за счет уравновешивания противоположных интересов различных видов, существующих вместе в рамках единого сообщества (например, хищник – жертва);
4) Сообщества основаны на количественной регуляции численности популяций одних видов другими;
5) Предельные размеры организма ограничены его внутренней наследственной программой, а размеры надорганизменной системы определяются внешними условиями.
Непосредственно живое окружение организма составляет его биоценотическую среду. Представители каждого вида способны существовать лишь в таком живом окружении, где связи с другими организмами обеспечивают им нормальные условия жизни. Иными словами, многообразные живые организмы встречаются на земле не в любом сочетании, а образуют определенные сожительства или сообщества, в которые входят виды, приспособленные к совместному обитанию. Приспособленность членов биоценоза к совместной жизни выражается в определенном сходстве требований к важнейшим абиотическим условиям среды и закономерных отношений друг с другом. Виды в биоценозе настолько приспособлены друг к другу, что их сообщество составляет системное целое; в природе не существует полезных и вредных организмов, все служит друг другу и взаимно приспособлено. Примеров взаимозависимости организмов очень много. Фитофаги не могут существовать без соответствующих растений, растения, опыляемые насекомыми, не могут размножаться в экосистеме, где нет необходимых для их опыления насекомых. В каждом биоценозе набор живых организмов является не случайным, а закономерным и любой биоценоз является устойчивой группировкой, повторяющейся во времени и пространстве.
Масштабы биоценотических группировок организмов очень разнообразны: от сообществ подушек лишайников на стволах деревьев или разлагающегося пня до населения целых ландшафтов - лесов, степей, пустынь и т.п.
Термин «биоценоз» в современной экологической литературе чаще употребляют применительно к населению территориальных участков, которые на суше выделяют по относительно однородной растительности (обычно по границам растительных ассоциаций), например, биоценоз ельника-кисличника, большого суходольного луга, сосняка-беломошника, большой ковыльной степи, пшеничного поля и т.д. При этом имеется в виду вся совокупность живых существ, растений, животных микроорганизмов, приспособленных к совместному обитанию на данной территории. В водной среде различают биоценозы, соответствующие экологическим подразделениям частей водоемов, например, больших прибрежных галечных, песчаных или илистых грунтов, абиссальных глубин, пелагических больших крупных водоворотов водных масс и т.п.
По отношению к более мелким сообществам (населению стволов или листвы деревьев, моховых кочек на болотах, нор, муравейников и т.д.) применяют разнообразные термины: «микросообщества», «биоценотические группировки», «биоценотические комплексы» и др.
Принципиальной разницы между биоценотическими группировками разных масштабов нет. Более мелкие сообщества входят составной, хотя и относительно автономной частью в более крупные, а те, в свою очередь, являются частями сообществ еще больших масштабов. Так, все живое население моховых и лишайниковых подушек на стволе дерева - это часть более крупного сообщества организмов, связанных с данным деревом и включающего его подкоровых и наствольных обитателей, население кроны, ризосферы и т.п.
В свою очередь эта группировка - лишь одна из составных частей лесного биоценоза. Последний входит в более сложные комплексы, образующие, в конечном счете, весь живой покров Земли. Таким образом, организация жизни на биоценотическом уровне иерархична. С увеличением масштабов сообществ усиливается их сложность и доля непрямых, косвенных связей между видами.
Биогеоценоз - исторически сложившаяся совокупность живых организмов (биоценоз) и абиотической среды вместе с занимаемым ими участком земельной поверхности (биотопом) (Рис. 8). Граница биогеоценоза устанавливается по границе растительного сообщества (фитоценоза) - важнейшего компонента биогеоценозов. Для каждого биогеоценоза характерен свой тип вещественно-энергетического обмена.
Итак, биогеоценоз - это составная часть природного ландшафта и элементарная биотерриториальная единица биосферы которая имеет упорядоченную структуру. Различают видовую, пространственную, трофическую и другие структуры биоценоза.
Разнообразие видов в биоценозе, соотношение их численности и биомассы всех популяций определяет его видовую структуру. Различают богатые и бедные видами биоценозы. В холодном климате тундры при большом дефиците тепла, в безводных жарких пустынях, в сильно загрязненных водоемах сообщества видов крайне скудные. В тех условиях, где факторы абиотической среды являются оптимальными для жизни, возникают чрезвычайно богатые видами сообщества. Видовой состав молодых формирующихся сообществ обычно беднее сложившихся, зрелых. Виды, преобладающие в биоценозе по численности, называются доминантными. Среди них выделяются те, которые свой жизнедеятельностью в наибольшей степени создают среду для всего сообщества и они называются эдификаторы. Основными эдификаторами биоценозов выступают определенные растения, но в некоторых случаях могут быть и животные.
Другим важным показателем биоценоза является размещение организмов в пространстве или пространственная структура. Пространственным границам биоценоза соответствует понятие биотоп – подразделение ландшафтной зоны, характеризующееся единым типом растительного покрова (фитоценоза). Именно фитоценоз определяет пространственные границы данной экологической системы. Горизонтальная структура биоценозов выражена их мозаичностью и реализуется в виде неравномерного распределения популяций отдельных видов по площади. Это определяется, с одной стороны, особенностями биотопов – неоднородностью почвенно-климатических условий, микроклимата и т.д., с другой стороны – взаимоотношениями отдельных видов как внутри популяции, так и между собой. На этой основе формируются разного рода группировки, в которых видовые популяции связаны между собой более тесными функциональными отношениями, чем с остальной частью биоценоза (в наземных биоценозах группировки автотрофов и гетеротрофов - концорции, в растительных сообществах - парцеллы).
Видовые популяции в составе биоценоза закономерно располагаются не только по площади, но и по вертикали в соответствии с биологическими особенностями каждого вида. Основную роль играет ярусность, т.е. размещение организмов в биоценозе по горизонтам почвы и по слоям воздушного пространства. Ярусная структура биоценоза создается растениями в наземных биоценозах. Ярусы бывают наземными и подземными, в водных экосистемах – подводные, надводные и др. В некоторых биоценозах ярусы выражены хорошо, в других слабо.
Функциональные взаимосвязи организмов в биоценозах многообразны (материальные, энергетические, химические, информационные). Кроме трофических связей, определяющих цепи питания (иногда очень своеобразных – паразитизм, симбиоз), существуют связи, основанные на том, что одни организмы становятся субстратами для других (топические связи), создают необходимый микроклимат и т.д. Можно проследить в биоценозе группы видов, связанные с определенным видом и полностью зависящие от последнего (концорции). Например, одной из концорций елового леса является популяция елей (детерминанты) с грибами, микориза которых оплетает корни, с бурундуками и белками, питающимися семенами (консорты). Детерминантами концорций являются популяции вида высшего растения, вокруг которого собираются сотни низших видов растений, грибов и животных, вступающих в различные взаимоотношения: мутуализм, комменсализм, паразитизм, трофические связи. Фабрические связи в биоценозах – связи, при которых особи одного вида используют особей другого вида ила частей тела для постройки необходимых им гнезд и убежищ. Таков, например, характер связи лесных птиц с лесными ценозами, предоставляющими им дупла или ветки для сооружения гнезд. Форические связи обеспечивают перенесение особей одного вида особями другого вида. Расселение и прорастание многих растений с сочными плодами зависит от присутствия животных, обеспечивающих перенесение их семян.
Биоценозы можно разделить на более мелкие, подчиненные единицы – мероцинозы, такие закономерно слагающиеся комплексы, которые зависят от биоценоза в целом. В годовом цикле изменяются численность, стадии развития и активности отдельных видов, что создает закономерные сезонные аспекты биоценозов.
Различают насыщенные и ненасыщенные биоценозы. В насыщенном биоценотическом сообществе все экологические ниши заняты и вселение нового вид невозможно без уничтожения или последующего вытеснения какого либо компонента. Ненасыщенные биоценозы характеризуются возможностью вселения новых видов без уничтожения существующих. Можно различать первичные биоценозы, сложившиеся без участия человека, (целинная степь, девственный лес) и вторичные, измененные деятельностью человека, (леса выросшие на месте вырубленных, водохранилища). Особую категорию составляют агробиоценозы, где компоненты сознательно регулируются человекам. Между первичными и агробиоценозами существует вся гамма переходов.
Биоценозы формируются в течение длительного промежутка времени и характеризуются определенной динамикой. Различают несколько стадий их формирования.
1. Колонии (пионерская группировка), например растения существуют поодиночке.
2. Группировка – биоценоз с незначительным количеством видов, территория (горизонтально и вертикально) не покрыта полностью.
3. Замкнутая группировка – территория покрыта полностью, но есть свободные ниши.
Основная функция биоценозов - поддержание круговорота веществ в биосфере – базируется на пищевых взаимоотношениях видов, что определяет трофическую структуру. Органические вещества, синтезированные автотрофными организмами, претерпевают многоэтапные химические превращения и в конечном итоге возвращаются в среду в виде минеральных веществ, вновь вовлекаемых в круговорот. Поэтому при всем многообразии видов входящих в биоценоз, обязательным является присутствие всех трех экологических групп организмов – продуцентов, консументов и редуцентов. Все виды разделяются на несколько групп в зависимости от места в общей системе круговорота веществ и потока энергии. Равнозначные по функциональному назначению виды образуют определенный трофический уровень, а взаимоотношения между видами разных уровней – систему цепей питания.
Прямые пищевые связи «растение – фитофаг - хищник – паразит» объединяют виды в цепи питания или трофические цепи. Каждый трофический уровень представлен не одним, а многими конкретными видами. Большинство животных использует в пищу более или менее широкий набор кормовых объектов. В результате помимо прямых «вертикальных» пищевых связей возникают боковые, объединяющие потоки вещества и энергии нескольких пищевых цепей. Таким путем возникают трофические сети, в которых множественность цепей питания выступает как приспособление к устойчивому существованию экосистемы в целом. Дублирование потоков вещества и энергии по большому числу параллельных трофических цепей поддерживает непрерывность круговорота при вероятных нарушениях отдельных пищевых цепочек.
Рассмотренные выше процессы связаны с синтезом и трансформацией органического вещества в трофических сетях. Процессы поэтапной деструкции и минерализации органических веществ выводятся в отдельный блок трофической структуры, называемой цепями разложения (детритными цепями). Выделение детритных цепей обусловлено тем, что минерализация органических веществ идет на всех трофических уровнях (растения и животные редуцируют органику до воды, диоксида углерода, минеральных солей и эти продукты выводятся в окружающую среду). Детритные цепи начинаются с разложения мертвого органического вещества особыми группами консументов – сапрофагами, которые механически, и отчасти химически разрушают его, подготавливая к воздействию редуцентов. В наземных экосистемах этот процесс сосредоточен преимущественно в подстилке и почве.
Положение популяции в составе экосистемы определяется с одной стороны, набором требований к абиотическим условиям, а с другой – комплексом связей с популяциями иных видов и формой участия в общих функциях биоценоза. Длительное существование многовидового сообщества привело к эволюционному становлению такой системы взаимоотношений, при которой каждый вид занимает определенное положение в составе биоценоза. Это функциональное и пространственное положение рассматривается как его экологическая ниша. Ее можно представить как объем многомерного пространства, который определен требованиями вида к экологическим факторам. Объем пространства, соответствующего требованиям вида к среде называется фундаментальной нишей, а реальное положение популяции в конкретной экосистеме – реализованной нишей. Реализованная ниша меньше фундаментальной т.к. в биоценозе часть подходящих для вида условий отсутствуют, либо этому мешают имеющиеся конкурентные взаимоотношения. Экологическую нишу можно определить как «профессию» вида в сообществе, что определяется физическим пространством, занимаемое популяцией, метом вида в системе внешних факторов и его функциональной ролью в экосистеме.
Основные формы межвидовых связей в биоценозах реализуются через сложные формы взаимодействия популяций различных видов в основе которых лежат трофические связи, обеспечивающие осуществление биологического круговорота. В общей форме можно выделить три основных вида межвидовых взаимоотношений: антибиоз, нейтрализм и симбиоз (Табл. 1). Антибиоз – крайнее выражение конкурентных отношений, при котором какой-то вид полностью препятствует поселению другой популяции в пределах зоны влияния. Антибиоз поддерживается главным образом химическим воздействием на потенциальных конкурентов и наиболее свойственен грибам и прокариотам. Ярким примером может быть деятельность ряда почвенных грибов (микромицетов), продуцирующих бактерицидные вещества. Некоторые из них стали основой производства антибиотиков.
Табл. 1. Типы взаимодействий между организмами в биоценозах
| Результаты взаимоотношений | Типы взаимоотношений | |
| + | + | Мутуализм - организмы получают относительно равную пользу |
| + | 0 | Комменсализм – «нахлебничество», присутствие комменсала для хозяина обычно не ощутимо |
| + | 0 | Квартиранство – использование хозяина для местообитания. |
| 0 | 0 | Нейтрализм – отсутствие взаимного влияния |
| Негативные взаимоотношения | ||
| - | - | Аллелопатия – взаимодействие между растениями, путем выделения продуктов жизнедеятельности (фитонцидов, антибиотиков) |
| - | - | Конкуренция – активное противодействие одного организма другому, борьба за существование |
| + | - | Хищничество – нападение одних животных на других и поедание их. Внутривидовое хищничество - каннибализм. |
| + | - | Паразитизм – жизнь организма-паразита за счет тканей и соков хозяина |
| 0 | - | Аменсализм – взаимодействие для одного вида безразлично, для другого - отрицательно |
Нейтрализм – тип отношений между видами, при котором они не формируют сколько-нибудь значительных прямых взаимодействия. Привести примеры таких взаимоотношений довольно сложно, т.к. многообразие биогеоцентических взаимоотношений приводит к тому, что большинство видов хотя бы косвенно влияют друг на друга. Чаще всего нейтрализм встречается в отношениях между видами, не принадлежащих к смежным трофическим уровням.
Симбиоз – система отношений, при которой формируются тесные функциональные взаимоотношения, выгодные для обеих видов (мутуализм) либо для одного из них (комменсализм). При выраженной форме мутуализма взаимная зависимость видов столь велика, что они не могут существовать изолировано. Например, жвачные животные (фитофаги) не могут усваивать клетчатку растений без кишечной микрофлоры.
Паразитизм – форма биотических взаимоотношений, при котором один вид получает преимущества за счет другого, которому наносится больший или меньший вред.
Наиболее важные преимущества живого организма как среды обитания обусловлено:
1. Постоянство внутренней среды организма создает высокую степень постоянства условий обитания. Это позволяет паразитам не вырабатывать многих механизмов, функция которых – приспособление к меняющимся условиям внешней среды.
2. Использование живого организма как среды обитания означает формирование «среды второго порядка» и упрощение взаимосвязей с внешней (первого порядка) средой. Биологические преимущества обитания в другом организме связаны с отсутствием необходимости сложных адаптаций.
3. Организм хозяина защищает паразита от потенциальных врагов.
4. Паразит питается за счет хозяина. Именно со взаимоотношений по линии питания и начиналось в большинстве случаев эволюция паразитизма.
В целом, общее биологическое преимущество, заложенное в основе эволюции паразитизма, заключается в возможности более экономного расхода энергии на различные (в первую очередь адаптивные) процессы, не связанные с прямой функцией поддержания жизни.
В масштабах биоценоза паразитарные связи объединяют ряд видов специфическими взаимоотношениями; на этой почве формируются внутриэкосистемные структурные образования, которые называют паразитарными системами или паразитоценозами. Паразитоценозом называют комплекс видов, поражающих общего хозяина; в это понятие включается как постоянные эндо- и эктопаразиты, так и временные (например, паразитоценоз норы грызунов). Паразитоценоз норы грызунов сходен с понятием концорции.
Динамика биоценозов.
Сложность межвидовых взаимоотношений, определяющих функциональную целостность биоценоза, отличается относительной свободой связей между отдельными компонетами. В зависимости от динамики численности и биологической активности отдельных популяций меняется направленность и интенсивность потоков вещества и энергии. Нестабильность абиотических условий определяет колебательный характер состава и функциональных связей в биоценозе. Динамичность – одно из фундаментальных свойств экосистем, отражающее не только зависимость от комплекса факторов, но и адаптивный ответ системы в целом на их воздействие.
При закономерных ритмичных суточных изменениях не происходит перестроек видового состава и основных форм взаимоотношений в биоценозе, поэтому говорят о суточных аспектах в данной экосистеме. Суточная динамика биоценозов отражается характером ритма жизнедеятельности видов, входящих в сообщество.
Сезонные изменения затрагивают видовой состав биоценозов (сезонные миграции, изменения растительного покрова) и функциональные связи (изменение трофических сетей).
Развивающееся единство различных видов, меняющееся в результате деятельности входящих в него компонентов, может приводить к закономерному изменению и смене биоценоза – сукцессии. Экологической сукцессией называется постепенное изменение структуры и состава биоценоза. Различают два основных типа сукцессий: вызываемые внутренними (эндогенными) и вызываемые внешними (экзогенными) факторами.
Пример эндогенной сукцессии – зарастание небольшого водоема. Отмирающие растения, ил откладываются на дне, уровень водоема постепенно поднимается. Растения, которые росли по берегам, постепенно распространяются к центру и занимают всю его площадь. Со временем пруд превращается в болото. Под влиянием внутренних факторов, в ходе сукцессии природа сама восстанавливает экологическое равновесие, которое было нарушено человеком. Растения, заселяющие поверхность шахтных терриконов (вначале травы, а затем кустарники и деревья) препятствуют выветриванию породы и загрязнению атмосферы.
Экзогенные сукцессии обусловлены нарушением экологического равновесия. Изменяя свой состав, биоценоз приспосабливается к новому фактору. Например, в результате сильного выпаса высокорослые травы исчезают. В ходе сукцессии им на смену приходят низкорослые растения с прижатыми к земле розетками листьев. Такие растения меньше страдают от копыт животных и их труднее скашивать.
Когда биоценоз достигает стабильной фазы развития, наступает климакс, который характеризуется особым распределением биогенных химических элементов. Они концентрируются в биомассе, в то время как абиогенная среда обеднена ими. Климаксные группировки отличаются тем, что в них больше детрита, а фитофаги отступают на вторую позицию по сравнению детритофагами. В зависимости от истории и длительности существования различают пионерские, сукцессионные и климаксные биоценозы.
Сукцессии как последовательный переход одного биоценоза в другой в пространстве и во времени сопровождающийся сменой состояний и свойств всех его компонентов, может возникнуть как под действием природных факторов, так и под действием человека. В связи с этим различают несколько форм сукцессии: антропогенную, пирогенную, катастрофическую и др.
Антропогенная сукцессия – это последовательная смена биогеоценозов, возникающая под влиянием хозяйственной деятельности человека, его прямым или косвенным влиянием на экосистему (вырубки леса, загазованность атмосферы, загрязнение сточными водами водоема).
Пирогенная сукцессия – это смена биоценозов в результате пожара, вне зависимости от их причины (природные или по вине человека).
Катастрофическая сукцессия – это с процесс, происходящий вследствие катастрофических для экосистемы происшествий: необычный паводок, массовое размножение вредителей, извержение вулкана и т.д.
Сукцессии в природе многообразны и разномасштабные. Иерархичность в организации сообществ проявляется в иерархичности сукцессионных процессов: более крупные преобразования биогеоценозов складываются из более мелких. Даже в стабильных экосистемах с хорошо отрегулированным круговоротом веществ постоянно осуществляется множество локальных сукцессионных смен, поддерживающих сложную внутреннюю структуру сообществ. По мере развития экосистемы число составляющих ее видов возрастает, а связи между ними становятся более сложными и разветвленными. Это приводит к более полному использованию ресурсов среды, к увеличению устойчивости природной системы. В конце концов, возникает устойчивая зрелая экосистема, находящаяся в равновесии со средой и способная сохраняться в течение длительного времени в относительно неизменном виде. Обычно в природе процесс сукцессии длится тысячи лет, но в отдельных случаях, например, после пожаров или при зарастании водоемов, можно наблюдать смену экосистем на глазах одного поколения людей. Несмотря на относительную устойчивость зрелых экосистем, они тоже могут заменяться другими. Это происходит, например, при резком изменении климата, а в последнее время в результате антропогенной деятельности (вырубка лесов, распашка земель, строительство городов и поселков, добыча полезных ископаемых и т.д.).
Особой разновидность биоценозов являются агроценозы, искусственно созданные человеком для своих целей с определенным уровнем и характером продуктивности. В настоящее время этими природными сообществами занято около 10% суши. Несмотря на то, что в агроценозе, как и в любой природной экосистеме, существуют обязательные трофические уровни – продуценты, консументы, редуценты, образующие типичные трофические связи, между этими сообществами существуют довольно большие различия:
1. В агроценозах резко снижено разнообразие организмов. Однообразие и видовую бедность агроценозов человек поддерживает специальной сложной системой агротехнических мер. На полях обычно культивируется один вид растений, в связи с чем резко обедняется и животное население, и состав микроорганизмов почвы. Однако даже самые обедненные агроценозы включают несколько десятков видов организмов, принадлежащих к разным систематическим группам. Например, в агроценоз пшеничного поля, кроме пшеницы, входят сорняки, насекомые-вредители пшеницы и хищники, беспозвоночные – обитатели надпочвенного слоя и почвы, разнообразные грибки и т. д.
2. Виды, культивируемые человеком, поддерживаются искусственным отбором и не могут выдерживать борьбу за существование без поддержки человека.
3. Агросистемы получают дополнительную энергию благодаря деятельности человека, обеспечивающего дополнительные условия роста культивируемых растений.
4. Чистая первичная продукция агроценоза (биомасса растений) удаляется из экосистемы в виде урожая и не поступает в цепи питания. Частичное потребление ее вредителями всячески пресекается деятельностью человека. В результате этого почва обедняется минеральными веществами, необходимыми для жизнедеятельности растений. Следовательно, снова необходимо вмешательство человека в виде внесения удобрений.
В агроценозах ослаблено действие естественного отбора и действует в основном искусственный отбор, направленный на максимальную продуктивность растений, нужных человеку, а не тех, которые лучше приспособлены к окружающим условиям. Таким образом, агроценозы, в отличие от природных систем, не являются саморегулирующимися, а регулируются человеком. Задачей такой регуляции является повышение продуктивности агроценоза. В отличие от природной экосистемы агроценоз неустойчив, он быстро разрушается, т.к. культурные растения не выдерживают конкуренции с дикорастущими и будут ими вытеснены.
Изучение биоценозов важно для рационального освоения земель и водных пространств, т.к. только правильное понимание регулятивных процессов в биосфере позволяет человеку изымать часть биопродукции без его нарушения и уничтожения. Все природные экосистемы связаны между собой, и вместе образуют живую оболочку Земли, которую можно рассматривать как самую большую экосистему, которая называется биосферой.
Вопросы для самоконтроля:
1. Характеристика биоценоза, его структура.
2. Характеристика пространственной и видовой структуры биоценоза.
3. Функциональные связи в биоценозах, их виды и характеристика.
4. Динамика формирования биоценозов, стадии этого процесса.
5. Понятие экологической ниши.
6. Типы взаимодействий между организмами.
7. Виды симбиотических взаимоотношений, их характеристика.
8. Виды негативных биотических взаимоотношений, их характеристика.
9. Динамика биоценозов. Сукцессии, их разновидности.
10. Агроценозы, их особенности и характеристика.
Учение об экосистеме
Понятие «экосистема» была введено английским ботаником А.Тенсли в 1935 году. Под экосистемой понимают стабильную природную систему, между живыми и неживыми компонентами которой постоянно происходит обмен веществ.
Экосистема - это любое сообщество живых существ вместе с его физической средой обитания, функционирующее как единое целое, объединенное вещественно-энергетическими и информационными взаимодействиями (Рис. 8).
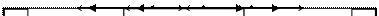
Рис. 8. Схема строения природной экосистемы.
Понятие экосистемы не ограничивается какими-то признаками ранга, размера, сложности или происхождения. Поэтому оно приложимо к относительно простым искусственным (аквариум, теплица, пшеничное поле, обитаемый космический корабль), так и к сложным естественным комплексам организмов и среды их обитания (озеро, лес, океан, биосфера).
Биотоп – участок земной поверхности (суши или водоема) с однотипными абиотическими условиями существования (микроклимат, рельеф, условия увлажнения и т.п.). Для каждого биотопа характерен специфический комплекс условий, который определяет видовой состав биоценоза (биотический компонент экосистемы). Биотоп является неорганическим компонентом экосистемы и общим местообитанием для всех членов сообщества. Члены сообщества так тесно взаимодействуют со средой обитания, что часто трудно рассматривать их отдельно.
Рассмотрение экосистемы важно в тех случаях, когда речь идет о потоках вещества и энергии, циркулирующих между живыми и неживыми компонентами природы, о динамике элементов, поддерживающих существование жизни, об эволюции сообществ. Ни отдельный организм, ни популяцию, ни сообщество в целом нельзя изучать в отрыве от окружающей среды. Экосистема, по сути, это то, что мы называем природой.
Экосистема - понятие очень широкое и применимо как к естественным (тундра, океан), так и к искусственным комплексам (аквариум). Поэтому для обозначения элементарной природной экосистемы экологи также используют термин "биогеоценоз". Экологические системы состоят их живых организмов и среды обитания – костной (атмосфера) и биокостной (почва, водоем и т.д.). Они иногда отделены друг от друга, но часто между ними имеются переходы. Примерами экологических систем являются озеро, лесной массив и т.д. От экосистем следует отличать биомы, под которыми понимаются значительные сообщества, приуроченные к определенным географическим зонам с их климатическими и почвенными зонами.
Экосистемы очень разнообразны (Рис. 9). Их состав зависит от многих факторов, в первую очередь от климата, геологических условий и влияния человека. Основные их типы - естественные и антропогенные.
Естественные экосистемы - формируются под влиянием природных факторов, хотя на них может оказывать влияние и человек. В естественном лесу человек заготавливает древесину и охотится, на степном пастбище пасет скот, в водоемах ловит рыбу. Он может влиять на экосистемы загрязняя атмосферу, почву, воду, однако влияние человека всегда меньше чем действие природных факторов.

Рис. 9. Классификация экосистем
Антропогенные экосистемы – создаются человеком в процессе хозяйственной деятельности. В их состав могут входить сохранившиеся более мелкие естественные экосистемы, различающиеся по источникам энергии, которые обеспечивают их жизнедеятельность.
Автотрофные сообщества находятся на энергетическом самообеспечении. Они разделяются на фотоавтотрофные – потребляющие солнечную энергию, которую усваивают растения, и хемоавтотрофные – использующие энергию окисления химических веществ (аммиака, соединений серы и т.д.). Фотоавтотрофными являются большинство экосистем, включая и сельскохозяйственные (агроэкосистемы). В последние человек вносит энергию удобрений, топлива сельскохозяйственной техники и т.д. Эту энергию называют антропогенной, ее роль незначительна по сравнению с потребляемой солнечной энергией.
Естественные хемотрофные экосистемы формируются в подземных водах. Антропогенные хемотрофные экосистемы человек создает в биологических очистных сооружениях для очистки сточных вод от загрязнения.
Гетеротрофные экосистемы получают энергию, уже накопленную разными организмами других экосистем или созданными человеком в результате органического и неорганического синтеза.
Примером естественной гетеротрофной экосистемы являются сообщества океанических глубин, куда не проникает солнечный свет. Живые организмы питаются за счет «питательного дождя», т.е. остатков организмов, которые падают на дно из освещенной солнцем автотрофной океанической системы.
Антропогенная гетеротрофная экосистема – город или промышленное предприятие. Энергия в эти системы поступает по линиям электропередач, по трубам нефтепроводов и газопроводов, в железнодорожных вагонах и на автотранспорте. Поступает в город сырье для работы предприятий и продукты питания для населения. Какое-то количество солнечной энергии фиксируется в этом сообществе, но она составляет небольшую часть от всей энергии потребляемой системой.
Наземные экосистемы объединяют в три основных типа:
1) экосистемы суши (степи, лиственные леса, северные хвойные леса, тундры, пустыни);
2) пресноводные экосистемы (проточных и непроточных водоемов);
3) морские экосистемы.
Схематическое, графическое изображение пищевой цепи получило название экологических пирамид. Различают три типа экологических пирамид: 1) чисел – учитывает численность отдельных организмов на каждом трофическом уровне; 2) биомассы – характеризует общую сухую или сырую массу организмов на разных уровнях; 3) энергии – показывает величину потока энергии (Рис. 10.).
Американский эколог Ю. Одум в предложенных пирамидах предположил, что исходной продукцией цепи является посев люцерны на площади 4 га, которой можно прокормить 4,5 телят (предполагается, что они едят только люцерну), а телятиной питается 12 летний мальчик (предполагается, что он потребляет только телятину). Расчеты, выполненные по приведенным показателям показали, что эффективность всех уровней невысока – люцерна использует 0,24 % всей солнечной энергии, поступившей на поле, телята – 8% энергии накопленной люцерной, и всего лишь 0,7% энергии, накопленной телятами используется на рост и развитие мальчика.
А.

Б.

В.

Рис. 10. Экологические пирамиды: А – чисел; Б. – биомассы; В. – энергии.
Дата: 2019-02-25, просмотров: 759.