Нуклеиновые кислоты, как и белки, необходимы для жизни. Они представляют собой генетический материал всех живых организмов вплоть до самых простых вирусов. Название «нуклеиновые кислоты» отражает тот факт, что локализуются они главным образом в ядре (nucleus — ядро). При специфическом окрашивании на нуклеиновые кислоты ядра бывают очень хорошо видны в световом микроскопе.
Выяснение структуры ДНК (дезоксирибонуклеиновой кислоты) — одного из двух существующих типов нуклеиновых кислот — открыло новую эпоху в биологии, так как позволило, наконец, понять, каким образом живые организмы хранят информацию, необходимую для регулирования их жизнедеятельности и каким образом передают эту информацию своему потомству. Нуклеиновые кислоты состоят из мономерных единиц, называемых нуклеотидами. Из нуклеотидов строятся чрезвычайно длинные молекулы — полинуклеотиды. Чтобы понять структуру полинуклеотидов, необходимо ознакомиться с тем, как построены нуклеотиды.
Молекула нуклеотида состоит из трех частей — пятиуглеродного сахара, азотистого основания и фосфорной кислоты.
Сахар. Сахар, входящий в состав нуклеотида, содержит пять углеродных атомов, т. е. представляет собой пентозу. Различают два вида сахаров входящих в состав нуклеиновых кислот это рибоза и дезоксирибоза. РНК содержат рибозу, а ДНК дезоксирибозу.
Основания. В обоих типах нуклеиновых кислот содержатся основания четырех разных видов: аденин (А), гуанин (Г), цитозин (Ц) и тимин (Т) или урацил (У) (соответственно в ДНК или РНК. Основания принято обозначать первой буквой их названия: А, Г, Т, У и Ц.
Фосфорная кислота. Нуклеиновые кислоты являются кислотами потому, что в их молекуле содержится фосфорная кислота.
Роль нуклеотидов в организме не ограничивается тем, что они служат строительными блоками нуклеиновых кислот; некоторые важные коферменты также представляют собой нуклеотиды. Таковы, например, аденозинтрифосфат (АТФ), циклический аденозинмонофосфат (цАМФ), кофермент А, никотинамидадениндинуклеотид (НАД), никотинамидадениндинуклеотидфосфат (НАДФ) и флавинадениндинуклеотид.
Два нуклеотида, соединяясь, образуют динуклеотид в результате реакции конденсации между фосфатной группой одного нуклеотида и сахаром другого. При синтезе полинуклеотидов этот процесс повторяется несколько миллионов раз. Таким путем строится неразветвленный сахарофосфатный остов полинуклеотида.
Нуклеиновые кислоты, подобно белкам, обладают первичной структурой (под которой подразу-мевается их нуклеотидная последовательность) и трехмерной структурой. Современную общепринятую теорию строения нуклеиновыхкислот разработали Джеймс Уотсон и Фрэнсис Крик (James Watson, Francis Crick) в Кавендишской лаборатории Кембриджского. Используя все физические и химические данные, какие оказались в их распоряжении, Уотсон и Крик стали строить пространственные модели ДНК в надежде на то, что рано или поздно им удастся получить достаточно убедительную структуру, согласующуюся со всеми этими данными.
Уотсон и Крик задались целью проверить предположение, что молекула ДНК состоит из двух спиральных полинуклеотидных цепей, удерживаемых вместе благодаря спариванию оснований, принадлежащих соседним цепям. Основания этих цепей соединяются в пары с помощью водородных связей, Аденин спаривается с тимином, а гуанин — с цитозином; АТ-пара соединяется двумя водородными связями, а ГЦ-пара — тремя. Уотсон увидел, что при таком сочетании основания оказываются очень точно подогнанными друг к другу и что общий размер и форма двух этих пар оснований одинаковы. Водородные связи при других сочетаниях оснований в принципе возможны, но они гораздо слабее.
Строение молекулы ДНК
Уотсон и Крик показали, что ДНК состоит из двух полинуклеотидных цепей. Каждая цепь закручена в спираль вправо, и обе они свиты вместе, т. е. закручены вправо вокруг одной и той же оси, образуя двойную спираль. Цепи антипараллельны, т. е. направлены в противоположные стороны. Каждая цепь состоит из сахарофосфатного остова, вдоль которого перпендикулярно длинной оси двойной спирали располагаются основания; находящиеся друг против друга основания двух противоположных цепей двойной спирали связаны между собой водородными связями. Вдоль оси молекулы соседние пары оснований располагаются на расстоянии 0,34 нм. одна от другой. Полный оборот спирали приходится на 3,4 нм, т. е. на 10 пар оснований. Никаких ограничений относительно последовательности нулеотидов в одной цепи не существует, но в силу правила спаривания оснований эта последовательность в одной цепи определяет собой последовательность нуклеотидов в другой цепи. Поэтому говорят, что две цепи двойной спирали комплементарны друг другу.
Уотсон и Крик опубликовали сообщение о своей модели ДНК в журнале «Nature» в 1953 г., а в 1962 г. они вместе с Морисом Уилкинсом были удостоены за эту работу Нобелевской премии
Когда Уотсон и Крик в 1953 г. заявили о спиральной структуре ДНК, они высказали также предположение, что генетическая информация, которая передается из поколения в поколение и контролирует жизнедеятельность клетки, заключена в молекуле ДНК в форме последовательности оснований. После того как было показано, что ДНК кодирует синтез белковых молекул, стало ясно, что последовательность оснований в ДНК должна кодировать последовательность аминокислот в белковых молекулах. Эта связь между основаниями и аминокислотами известна под названием генетического кода. А элементарную единицу генетического кода определяющюю структурные, физиологические и биохимические признаки организма, было предложено назвать ген.
Ген было принято определять как наименьший участок хромосомы, обусловливающий синтез определенного продукта. Теперь мы знаем, что гены кодируют синтез белков. Поэтому ген можно определить, как участок ДНК, кодирующий определенный белок. Это определение можно еще больше уточнить, назвав ген участком ДНК, кодирующим определенный полипептид, поскольку некоторые белки состоят не из одной, а из нескольких полипептидных цепей, а поэтому кодируются более, чем одним геном
Как говорилось выше молекула ДНК построена из оснований четырех типов: аденина (А), гуанина (Г), тимина (Т) и цитозина (Ц). Каждое основание составляет часть нуклеотида, а нуклеотиды соединены в полинуклеотидную цепь; они обозначаются начальными буквами своих названий. И четыре буквы «алфавита» позволяют записывать инструкции для синтеза потенциально бесконечного числа различных белковых молекул. Существуют 20 аминокислот, из которых построены белки и которые должны кодироваться основаниями, входящими в состав ДНК. Если бы положение одной аминокислоты в первичной структуре какого-либо белка определяло одно основание, то этот белок мог бы содержать только четыре различные аминокислоты. Если бы каждая аминокислота кодировалась двумя основаниями, то с помощью такого кода можно было бы определить 16 аминокислот. Включение в белковые молекулы всех 20 аминокислот может обеспечить только код, состоящий из трех оснований. Такой код может давать 64 сочетания оснований, что более чем достаточно. Поэтому Уотсон и Крик предсказали, что код должен быть триплетным.
Структура РНК
РНК содержится во всех живых клетках в виде одноцепочечных молекул. Она отличается от ДНК тем, что содержит в качестве пентозы рибозу (вместо дезоксирибозы), а в качестве одного из пиримидиновых оснований — урацил (вместо тимина). Анализ РНК, содержащейся в клетках, показал, что существуют три типа РНК, участвующих в синтезе белковых молекул. Это матричная, или информационная, РНК (мРНК), транспортная РНК (тРНК) и рибосомная РНК (рРНК). Все три РНК синтезируются непосредственно на ДНК, а количество РНК в каждой клетке находится в прямой зависимости от количества вырабатываемого этой клеткой белка.
Матричная РНК
Как показали анализы, мРНК составляет 3—5% всей РНК клетки. Это одноцепочечная молекула, образующаяся на одной из цепей ДНК в процессе, называемом транскрипцией. При синтезе мРНК копируется только одна цепь молекулы ДНК. Последовательность оснований в мРНК представляет собой комплементарную копию соответствующей цепи ДНК; длина ее варьирует в зависимости от длины полипептидной цепи, которую она кодирует. Большая часть мРНК находится в клетке лишь короткое время. У бактерий это может быть всего несколько минут, тогда как в развивающихся эритроцитах мРНК может служить матрицей для синтеза гемоглобина в течение нескольких дней.
Рибосомная РНК
Рибосомная РНК составляет примерно 80% всей РНК клетки. Она кодируется генами, находящимися в ДНК нескольких хромосом, расположенных в участке ядрышка, известном под названием ядрышкового организатора. Последовательность оснований в рРНК сходна у всех организмов — от бактерий до высших растений и животных. рРНК содержится в цитоплазме, где она связана с белковыми молекулами, образуя вместе с ними клеточные органеллы, называемые рибосомами.
На рибосомах происходит синтез белка. Здесь «код», заключенный в мРНК, транслируется в аминокислотную последовательность полипептидной цепи.
Транспортная РНК.
Существование транспортной РНК (тРНК) было постулировано Криком. У каждой аминокислоты имеется собственная семья молекул тРНК. Они доставляют содержащиеся в цитоплазме аминокислоты к рибосоме. Таким образом, тРНК служит промежуточной молекулой между триплетным кодом в мРНК и аминокислотной последовательностью полипептидной цепи. На долю тРНК приходится примерно 15% всей клеточной РНК; у этих РНК самая короткая полинуклеотидная цепь — в нее входит в среднем 80 нуклеотидов. В каждой отдельной клетке содержится более 20 различных молекул тРНК. Последовательность оснований в триплете антикодона строго соответствует той аминокислоте, которую переносит данная молекула тРНК. Каждая аминокислота присоединяется к одной из своих специфичных тРНК при участии фермента аминоацил-тРНК-синтетазы. В результате образуется комплекс аминокислота-тРНК, известный как аминоацил-тРНК, который способен образовывать связь между аминокислотами. Таким образом синтезируется полипептидная цепь.
В результате установления взаимосвязи между ДНК и наследственными процессами, у живых организмов, возникла наука генетика. Ключевым понятием современной генетики является понятие «наследственность» - универсальное свойство живых организмов обеспечивать материальную и функциональную преемственность между поколениями. Наследование (процесс) следует отличать от наследственности (свойства). Наследование - процесс передачи информации о признаках.
В основе наследования лежат процессы удвоения, распределения и объединения генетического материала, которыми во многом обусловливаются закономерности наследования.
Функционально неделимой элементарной единицей наследственности является ген. Полное описание структуры генов какого-либо организма подразумевает описание последовательности нуклеотидов в ДНК этого организма. Однако описание последовательности нуклеотидов в ДНК даже мельчайших вирусов составляет колоссальную проблему, практически неразрешимую для ДНК высших организмов. Геном кишечной палочки Escherichia coli состоит примерно из 3,2x106 нуклеотидных пар. Для каждой нуклеотидной пары существуют четыре возможности комбинации (АТ, ТА, ГЦ, ЦГ), поэтому число возможных нуклеотидных последовательностей в генотипе Е. coli составляет 4 3,2x106 В генотипе человека их намного больше.
Рибосомные РНК содержат 3000–5000 нуклеотидов; молекулярная масса — 1 000 000–1 500 000. На долю рРНК приходится 80–85% от общего содержания РНК в клетке. В комплексе с рибосомными белками рРНК образует рибосомы — органоиды, осуществляющие синтез белка. В эукариотических клетках синтез рРНК происходит в ядрышках. Функции рРНК: 1) необходимый структурный компонент рибосом и, таким образом, обеспечение функционирования рибосом; 2) обеспечение взаимодействия рибосомы и тРНК; 3) первоначальное связывание рибосомы и кодона-инициатора иРНК и определение рамки считывания, 4) формирование активного центра рибосомы.
Информационные РНК разнообразны по содержанию нуклеотидов и молекулярной массе (от 50 000 до 4 000 000). На долю иРНК приходится до 5% от общего содержания РНК в клетке. Функции иРНК: 1) перенос генетической информации от ДНК к рибосомам, 2) матрица для синтеза молекулы белка, 3) определение аминокислотной последовательности первичной структуры белковой молекулы.
Вопрос 5. Строение хромосом.
Хромосомы (термин «хромосома» предложил В. Вальдейер в 1888 году) представляют собой структуры клеточного ядра, которым свойственны способность к самовоспроизведению и сохранению индивидуальных черт строения (структурной и функциональной индивидуальности) в ряду поколений. Размещение генов в хромосомах влияет на соотносительное наследование информации о признаках.
Как показывает анализ хромосом эукариотических клеток, они состоят из дезоксирибонуклеиновой кислоты (ДНК) и белка, а также небольшого количества хромосомной РНК. («Хромосомы» прокариотических клеток — бактерий — состоят из одной ДНК.) Молекула ДНК несет отрицательные заряды, распределенные по всей ее длине, а присоединенные к ней белковые молекулы — гистоны — заряжены положительно. Этот комплекс ДНК—белок называется хроматином.
Большое количество ДНК, содержащейся в клетках, сопряжено с проблемой упаковки. Например, одна клетка человека содержит примерно 2,2 м ДНК, распределенной между 46 хромосомами. Каждая хромосома, таким образом, содержит около 4,8 см (48 000 мкм) ДНК. Длина хромосом человека составляет в среднем 6 мкм, коэффициент упаковки 8000:1. Такая концентрация достигается благодаря компактной упаковке нуклеиновой кислоты.
Выделяют следующие уровни компактизации ДНК.
1. Нуклеосомный уровень. Нуклеосома - это ДНК - гистоновый комплекс, который выглядит как частица дисковидной формы диаметром 11 нм. Впервые нуклеосомы были описаны в 1974г. А. Олинс и Д. Олинс. Каждая нуклеосома состоит из белкового кора или октамера и 2 оборотов фрагмента двухцепочечной ДНК. Нуклеосомы связываются участками ДНК (линкерная ДНК) свободными от контакта с белковым кором. Визуально она представляет собой структуру, напоминающую бусы на нитке, где в качестве бусин выступают нуклеосомы, а в качестве нитки – цепь ДНК
Укладка линкерного участка ДНК (60-80 п.н.) и соединение нуклеосом друг с другом идут с помощью гистона Н1. Молекула этого белка имеет центральную (глобулярную) часть и вытянутые «плечи». Центральная часть прикрепляется к специфическому участку на поверхности кора, вытянутые «плечи» соединяют соседние нуклеосомы. При этом ДНК наматывается на соседние коры каждый paз в противоположном направлении. Нуклеосомный уровень повышает плотность упаковки ДНК в 7-10 раз.
2.Нуклеомерный уровень. Дальнейшая компактизация ДНК в составе хроматина связана с образованием нуклеосомных комплексов. Образуется компактная хроматиновая фибрилла построенная либо по типу соленоида (спиральный тип укладки), либо по нуклеомерному типу (4-12 нуклеосом образуют глобулу). Нуклеомерная укладка хроматина способствует укорочению нити ДНК примерно в 6 раз, а оба уровня приводят к компактизации ДНК в среднем в 50 раз.
3. Хромомерный уровень. Следующий этап компактизации ДНК связан с образованием петлеобразных структур, которые называются хромомерами. Средняя длина петли (300-400 нм.) сходна у различных организмов (дрозофила и человек) и включает примерно 50 тысяч оснований. Такую петельную структуру называют интерфазной хромонемой. Негистоновые белки образуют ось, или осевой скелет, – непрерывный тяж, к которому крепятся петли ДНП, имеющие форму хроматиновой фибриллы. На петельном уровне ДНК может достаточно легко освобождаться от упаковывающих ее белков, и на соответствующих участках становится возможной транскрипция (то есть синтез РНК). Укорочение фибриллы на этом уровне происходит в среднем 25 раз, а на всех 3 уровнях в 1000-1500 раз.
4. Хромонемный уровень. При делении клеток идет дальнейшая компактизация хромосом - образование более крупных петель из хромомерной фибриллы. Образуется путем конденсации (укладки) петельных фибрилл в отдельные участки – хромомерные (утолщенные) центры, которые у некоторых видов эукариот выглядят как узелки. При этом в самом конце интерфазы образуется серия динамических петель с большой толщиной (шириной). В итоге достигается еще более плотная упаковка хроматина, прежде всего цепи ДНК. На поверхности упакованные молекулы ДНК несут множество белков, которые образуют подобие чехла.
5. Хромосомный уровень. Дальнейшая компактизация хромосом обеспечивается петельной укладкой хромонемной нити, что сокращает их длину примерно в 10 раз. На этом этапе происходит объединение петель имеющих одинаковую организацию, образуются блоки или минидиски. В образовании одного минидиска участвуют примерно около 20 петель. Таким образом, за счет нескольких уровней компактизации длина ДНК сокращается примерно в 10000 раз. Конденсация хромосом из деконденсированного состояния - это не спирализация, а очень сложный комплекс компактизации, связанный не только с изменением их линейных размеров, но и с регуляцией их работы в процессе жизнедеятельности клетки.
Кроме того, компактизация хромосомы - важнейший процесс, связанный с точной передачей наследственной информации очередному поколению.
Метафазная хромосома (хромосомы изучаются в метафазу митоза) состоит из двух хроматид. Любая хромосома имеет первичную перетяжку (центромеру), которая делит хромосому на плечи. Центромера определяет движение хромосомы во время клеточного деления. Некоторые хромосомы имеют вторичную перетяжку и спутник. Спутник — участок короткого плеча, отделяемый вторичной перетяжкой. Хромосомы, имеющие спутник, называются спутничными. Концы хромосом называются теломерами. В зависимости от положения центромеры выделяют: а) метацентрические (равноплечие), б) субметацентрические (умеренно неравноплечие), в) акроцентрические (резко неравноплечие) хромосомы.
Соматические клетки содержат диплоидный (двойной — 2n) набор хромосом, половые клетки — гаплоидный (одинарный — n). Диплоидный набор аскариды равен 2, дрозофилы — 8, шимпанзе — 48, речного рака — 196. Хромосомы диплоидного набора разбиваются на пары; хромосомы одной пары имеют одинаковое строение, размеры, набор генов и называются гомологичными.
Кариотип — совокупность сведений о числе, размерах и строении метафазных хромосом. У представителей разных видов кариотипы разные, одного вида — одинаковые. Аутосомы — хромосомы, одинаковые для мужского и женского кариотипов. Половые хромосомы — хромосомы, по которым мужской кариотип отличается от женского.
Вопрос 6. Биосинтез белка .
Биосинтез белков является важнейшим процессом анаболизма. Все признаки, свойства и функции клеток и организмов определяются в конечном итоге белками. Белки недолговечны, время их существования ограничено. В каждой клетке постоянно синтезируются тысячи различных белковых молекул. В начале 50-х гг. ХХ в. Ф. Крик сформулировал центральную догму молекулярной биологии: ДНК → РНК → белок. Согласно этой догме способность клетки синтезировать определенные белки закреплена наследственно, информация о последовательности аминокислот в белковой молекуле закодирована в виде последовательности нуклеотидов ДНК. Участок ДНК, несущий информацию о первичной структуре конкретного белка, называется геном. Гены не только хранят информацию о последовательности аминокислот в полипептидной цепочке, но и кодируют некоторые виды РНК: рРНК, входящие в состав рибосом, и тРНК, отвечающие за транспорт аминокислот. В процессе биосинтеза белка выделяют два основных этапа: транскрипция — синтез РНК на матрице ДНК (гена) — и трансляция — синтез полипептидной цепи.
В биосинтезе белка активное участие принимают различные молекулы РНК, рибосомы и мономерные молекулы – разные аминокислоты, из которых строится полимерная молекула – протеин, или белок.
План строения белка закодирован в молекуле ДНК, но сама она в синтезе белковых молекул участия не принимает, а служит лишь матрицей для синтеза информационной (или матричной) РНК (иРНК, или мРНК). Поэтому процесс синтеза белка складывается из двух этапов: создания иРНК и сборки молекулы белка по информации, заключенной в молекуле иРНК.
Первый этап – создание иРНК, которая путем «списывания» копирует последовательности нуклеотидов с определенных участков ДНК. Данный этап получил название транскрипция (от лат. transcriptio – переписывание).
Второй этап – этап непосредственного создания молекул белка – называют трансляцией (от лат. translatio – передача, перенесение).
Также следует обратить внимание на то, что появившийся таким путем первый транскрипт РНК еще не является готовой иРНК. Списанные копии несут в себе полную информацию, включая и интроны(участки не кодирующие ген), и экзоны(соответственно несущие генетическую информацию. Только после процесса созревания (сплайсинга, процессинга) первоначально списанный транскрипт РНК (преРНК) превратится в «зрелую» иРНК. Затем иРНК выходит из ядра в цитоплазму и, соединившись с рибосомой, начинает трансляцию – синтез белка из аминокислот с участием рибосом, ферментов, рРНК и тРНК.
Транскрипция у бактерий
Совсем иначе синтез иРНК осуществляется у прокариотических клеток (бактерий) и вирусов-бактериофагов. На кольцевой молекуле бактериальной ДНК (бакДНК) участки генетического материала, с которого осуществляется транскрипция одной иРНК, представлены нуклеотидными комплексами – оперонами (от лат. operor – работаю, действую). Оперон считают основной единицей генетического материала на уровне транскрипции у бактерий.
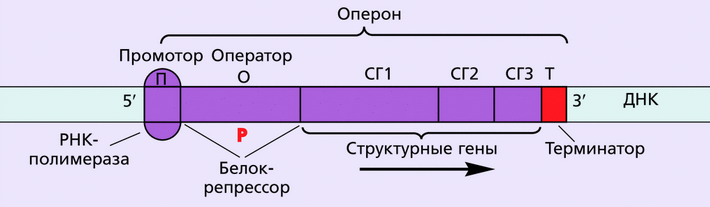 Структура оперона
Структура оперона
Наиболее важным компонентом оперонной регуляции синтеза РНК являются ген-регулятор, кодирующий белок-репрессор (подавитель), и оператор, к которому имеет сродство репрессор. Оператор и тесно сцепленные с ним структурные гены (один, два или несколько), находящиеся под его контролем, образуют оперон. Ген-регулятор не входит в состав оперона, хотя и может быть сцеплен с ним.
Каждый оперон включает в себя особые регуляторные зоны: участок начала транскрипции – промотор – и находящийся рядом с ним оператор, сцепленный со структурными генами; терминатор, сигнализирующий о прекращении транскрипции, расположен в конце оперона.
Промотор представляет собой короткую последовательность нуклеотидов ДНК (несколько десятков), с которым связывается РНК-полимераза, осуществляющая транскрипцию ДНК. Присоединившись к промотору, РНК-полимераза разрывает водородные связи между цепями ДНК, раскручивает спираль. В итоге на этом участке образуются две раздельные цепи ДНК, на одной из которых идет транскрипция, обеспечивающая синтез иРНК. Присоединение к иРНК новых нуклеотидов идет только на 3'-конце, поэтому синтезируемая цепь иРНК удлиняется в направлении 5' → 3'. Удлинение иРНК совершается до тех пор, пока не дойдет до терминатора. Здесь транскрипция прекращается, РНК-полимераза отделяется от матрицы и от синтезированной цепи иРНК. По завершении списывания РНК-полимераза вновь готова начать процесс транскрипции. Напомним, что созревания молекул РНК у прокариотических клеток не происходит, поэтому синтезируется сразу иРНК.
Однако процесс списывания информации с ДНК происходит лишь в том случае, если оператор не связан с репрессором. Если эта связь имеется, РНК-полимераза не может двигаться вдоль оперона. Пока репрессор связан с оператором, транскрипция не происходит, она блокирована репрессором. Для разблокирования репрессора необходимо, чтобы к промотору был присоединен активатор – белок позитивного (положительного) контроля. С момента присоединения белка-активатора к промотору репрессор блокируется и РНК-полимераза начинает транскрипцию.
Ген — участок молекулы ДНК, кодирующий первичную последовательность аминокислот в полипептиде или последовательность нуклеотидов в молекулах транспортных и рибосомных РНК. ДНК одной хромосомы может содержать несколько тысяч генов, которые располагаются в линейном порядке. Место гена в определенном участке хромосомы называется локусом. Особенностями строения гена эукариот являются: 1) наличие достаточно большого количества регуляторных блоков, 2) мозаичность (чередование кодирующих участков с некодирующими). Экзоны (Э) — участки гена, несущие информацию о строении полипептида. Интроны (И) — участки гена, не несущие информацию о строении полипептида. Число экзонов и интронов различных генов разное; экзоны чередуются с интронами, общая длина последних может превышать длину экзонов в два и более раз. Перед первым экзоном и после последнего экзона находятся нуклеотидные последовательности, называемые соответственно лидерной (ЛП) и трейлерной последовательностью (ТП). Лидерная и трейлерная последовательности, экзоны и интроны образуют единицу транскрипции. Промотор (П) — участок гена, к которому присоединяется фермент РНК-полимераза, представляет собой особое сочетание нуклеотидов. Перед единицей транскрипции, после нее, иногда в интронах находятся регуляторные элементы (РЭ), к которым относятся энхансеры и сайленсеры. Энхансеры ускоряют транскрипцию, сайленсеры тормозят ее.
Транскрипция у эукариот
Цепь иРНК синтезируется непосредственно на одной из цепей молекулы ДНК, которая в этом процессе служит матрицей. Синтезируемая цепь иРНК при этом точно копирует нуклеотидную последовательность матрицы по принципу комплементарности, с той лишь разницей, что тимину (Т) в ДНК соответствует урацил (У) в РНК. Весь процесс синтеза иРНК совершается с помощью особого фермента – РНК-полимеразы, передвигающегося вдоль цепи молекулы ДНК. Сама ДНК в этом процессе не изменяется и не расходуется, сохраняется в прежнем виде и потому сразу готова к новому списыванию с нее информации на иРНК.
На протяжении всей молекулы ДНК таких участков списывания информации может быть одновременно очень много. Многократным списыванием информации с каждого участка создается тот поток различных копий иРНК с ДНК, который обеспечивает программирование биосинтеза всех необходимых белков клетки.
Транскрипция — синтез РНК на матрице ДНК. Осуществляется ферментом РНК-полимеразой.
РНК-полимераза может присоединиться только к промотору, который находится на 3'-конце матричной цепи ДНК, и двигаться по этой матричной цепи ДНК. Синтез РНК происходит на одной из двух цепочек ДНК в соответствии с принципами комплементарности и антипараллельности. Строительным материалом и источником энергии для транскрипции являются рибонуклеозидтрифосфаты (АТФ, УТФ, ГТФ, ЦТФ).
В результате транскрипции образуется «незрелая» иРНК (про-иРНК), которая проходит стадию созревания или процессинга. Процессинг включает в себя: 1) КЭПирование 5'-конца т.е. достраивание с образованием особой структуры, ответственной за последующее связывание молекулы мРНК с рибосомой, 2) полиаденилирование 3'-конца (присоединение нескольких десятков адениловых нуклеотидов), 3) сплайсинг (вырезание интронов и сшивание экзонов). В зрелой иРНК выделяют КЭП, транслируемую область (сшитые в одно целое экзоны), нетранслируемые области (НТО) и полиадениловый «хвост».
Транслируемая область начинается кодоном-инициатором, заканчивается кодонами-терминаторами. НТО содержат информацию, определяющую поведение РНК в клетке: срок «жизни», активность, локализацию.
Транскрипция и процессинг происходят в клеточном ядре. Зрелая иРНК приобретает определенную пространственную конформацию, окружается белками и в таком виде через ядерные поры транспортируется к рибосомам; иРНК эукариот, как правило, моноцистронны (кодируют только одну полипептидную цепь).
Трансляция
Трансляция — синтез полипептидной цепи на матрице иРНК.
Органоиды, обеспечивающие трансляцию, — рибосомы. У эукариот рибосомы находятся в некоторых органоидах — митохондриях и пластидах, в свободном виде в цитоплазме и на мембранах эндоплазматической сети. Таким образом, синтез белковых молекул может происходить в цитоплазме, на шероховатой эндоплазматической сети, в митохондриях и пластидах. В цитоплазме синтезируются белки для собственных нужд клетки; белки, синтезируемые на ЭПС, транспортируются по ее каналам в комплекс Гольджи и выводятся из клетки. В рибосоме выделяют малую и большую субъединицы. Малая субъединица рибосомы отвечает за генетические, декодирующие функции; большая — за биохимические, ферментативные.
В малой субъединице рибосомы расположен функциональный центр (ФЦР) с двумя участками — пептидильным (Р-участок) и аминоацильным (А-участок). В ФЦР может находиться шесть нуклеотидов иРНК, три — в пептидильном и три — в аминоацильном участках.
Для транспорта аминокислот к рибосомам используются транспортные РНК, тРНК. Длина тРНК от 75 до 95 нуклеотидных остатков. Они имеют третичную структуру, по форме напоминающую лист клевера. В тРНК различают антикодоновую петлю и акцепторный участок. В антикодоновой петле РНК имеется антикодон, комплементарный кодовому триплету определенной аминокислоты, а акцепторный участок на 3'-конце способен с помощью фермента аминоацил-тРНК-синтетазы присоединять именно эту аминокислоту (с затратой АТФ). Таким образом, у каждой аминокислоты есть свои тРНК и свои ферменты, присоединяющие аминокислоту к тРНК.
теоретически может быть 61 вид тРНК с соответствующими антикодонами. Но кодируемых аминокислот всего 20 видов, значит, у одной аминокислоты может быть несколько тРНК. Установлено существование нескольких тРНК, способных связываться с одним и тем же кодоном (последний нуклеотид в антикодоне тРНК не всегда важен), поэтому в клетке обнаружено всего около 40 различных тРНК.
Синтез белка начинается с того момента, когда к 5'-концу иРНК присоединяется малая субъединица рибосомы, в Р-участок которой заходит метиониновая тРНК (транспортирующая аминокислоту метионин). Следует отметить, что любая полипептидная цепь на N-конце сначала имеет метионин, который в дальнейшем чаще всего отщепляется. Затем происходит присоединение большой субъединицы рибосомы, и в А-участок поступает вторая тРНК, чей антикодон комплементарно спаривается с кодоном иРНК, находящимся в А-участке.
Пептидилтрансферазный центр большой субъединицы катализирует образование пептидной связи между метионином и второй аминокислотой. Отдельного фермента, катализирующего образование пептидных связей, не существует. Энергия для образования пептидной связи поставляется за счет гидролиза ГТФ.
Как только образовалась пептидная связь, метиониновая тРНК отсоединяется от метионина, а рибосома передвигается на следующий кодовый триплет иРНК, который оказывается в А-участке рибосомы, а метиониновая тРНК выталкивается в цитоплазму. На один цикл расходуется 2 молекулы ГТФ. В А-участок заходит третья тРНК, и образуется пептидная связь между второй и третьей аминокислотами.
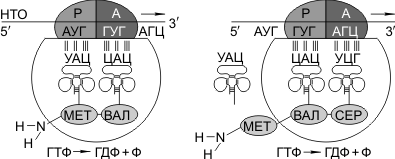
Трансляция идет до тех пор, пока в А-участок не попадает кодон-терминатор (УАА, УАГ или УГА), с которым связывается особый белковый фактор освобождения. Полипептидная цепь отделяется от тРНК и покидает рибосому. Происходит диссоциация, разъединение субъединиц рибосомы.
Скорость передвижения рибосомы по иРНК — 5–6 триплетов в секунду, на синтез белковой молекулы, состоящей из сотен аминокислотных остатков, клетке требуется несколько минут.
В трансляции можно выделить три стадии: а) инициации (образование иницаторного комплекса), б) элонгации (непосредственно «конвейер», соединение аминокислот друг с другом), в) терминации (образование терминирующего комплекса).
Дата: 2019-02-19, просмотров: 378.