Однако верующий дарвинист возразит Вам, что строгих установленных примеров эволюции под действием естественного отбора у него нет не потому, что подобной эволюции, на самом деле, не существует в природе. А просто потому, что эволюция под действием естественного отбора – это длительный процесс, который по этой причине пронаблюдать и строго зафиксировать – почти невозможно.
Практика показывает, что такое возражение дарвинист озвучивает примерно в 80% случаев. Правда, только после того, как уже выяснится, что со строгими примерами дарвиновской эволюции в биологии – полный «швах». А до выяснения этого обстоятельства, дарвинист будет уверять Вас в том, что теория естественного отбора:
- Давно и неопровержимо доказана наукой на огромном числе установленных примеров.
И только после того, как выяснится, что «огромное число установленных примеров», на самом деле, является жалкой горсткой из десятка научных публикаций, половина из которых уже раскритикована, а другая половина была сотни раз переписана из одного учебника в другой, и в результате эти «установленные примеры» встречаешь уже как старых, но изрядно надоевших друзей… вот только после этого дарвинист начнёт оправдываться, что столь жалкая кучка «установленных примеров» обусловлена «большой длительностью эволюционных процессов».
В качестве наглядного примера приведу прямую цитату из оправдательной речи очередного верующего дарвиниста, которую этот дарвинист произнёс после того, как у него (очень быстро) закончились «примеры установленной эволюции»:
- Вы противоречите самому себе, вы просите нечто, что наблюдалось за короткое время – теория эволюции требует гораздо большее время, т.е. в качестве доказательства вы хотите опровержение теории эволюции.
На подобные речи следует отвечать таким образом.
Прежде всего, необходимо напомнить верующему дарвинисту, что человек, который не может привести для обоснования своей веры никаких прямых доказательств, должен вести себя скромнее:
- Нет, что Вы. Это не я противоречу себе. Это верующие дарвинисты противоречат сами себе. Им надо определиться - или они признаются, что прямых доказательств у теории естественного отбора - нет (и допустим, быть не может, потому что для этого нужно много времени)... но тогда они должны принять более скромный вид, и не заявлять, что «эволюция - это факт», во всяком случае, по дарвиновскому механизму. Или же, если они всё-таки претендуют на то, что «Дарвин - это отец всех народов» (в биологии), тогда они обязаны предъявить серьезные доказательства этого (прямые эмпирические факты).
После этого Вы должны развенчать миф о том, что эволюционные изменения по механизму естественного отбора требуют много времени. Потому что на самом деле это не так. В очень многих случаях эволюционные изменения могут быть достаточно быстрыми, чтобы биологи могли их зафиксировать. Компьютерное моделирование показывает - если какой-то признак повышает выживаемость особей на 10%, то насыщение популяции особями, обладающими этим признаком, должно произойти всего примерно за 150 поколений.[91]
А 150 поколений - это для огромного числа живых существ - всего 150 лет. А для очень многих живых существ (даже для некоторых позвоночных животных) всего 30-50 лет. В большом числе случаев – даже еще меньше. Многие тропические насекомые могут производить до 10 поколений в год. Например, красивая тропическая бабочка Hypolimnas misippus имеет жизненный цикл длиной всего 23 дня. Что позволяет ей производить 6-7 поколений за один сезон размножения с августа по февраль (Atluri et al., 2013). Таким образом, 150 поколений для этой бабочки могут пролететь всего за 20-25 лет даже с учетом сезонного характера её размножения, то есть, несмотря на перерывы в размножении в течение года. И подобных насекомых в природе с избытком.
Обязательно напомните верующему дарвинисту про знаменитый пример с «эволюцией березовых пядениц». Как мы все знаем, «эволюция березовых пядениц» произошла в лесах Англии практически на наших глазах. В промышленно загрязненных районах Англии светлая форма этой бабочки была почти вытеснена появившейся темной формой. И на это хватило всего примерно 50 лет.
А потом, после того как промышленное загрязнение в этих районах уменьшилось, насекомоядные птицы опять взялись за естественный отбор березовых пядениц, но уже в обратном направлении – в сторону опять светлой формы. И на этот отбор тоже хватило всего нескольких десятков лет. Причем аналогичную картину (и аналогичную скорость естественного отбора) биологи зафиксировали не только в Англии, но еще и в Америке. Несмотря на то, что березовая пяденица в этих регионах даёт только одно поколение в год.
Вот 50 лет – это как раз и есть примерно ожидаемая скорость замены в популяциях вредного признака на полезный при том давлении естественного отбора, который был установлен для березовых пядениц. Как известно, в полевых экспериментах Kettlewell (Kettlewell, 1955; Kettlewell, 1956) разница в выживаемости разных цветовых форм березовых пядениц в природе составила (в разных экспериментах) от 17% (за один день) до вообще двойного преимущества (в экспериментах по выпуску и возвращению бабочек). А в более поздних исследованиях - примерно 10% (Cook et al., 2012). В детальных отчетах второй половины двадцатого века, которые регистрировали обратное снижение частоты встречаемости темной формы, было установлено, что вместе с падением уровня загрязнения, частота темной формы березовой пяденицы снизилась с 90% до менее 10% всего за 40 лет. Установив озвученное различие в выживаемости разных цветовых форм этих бабочек в природе, и озвученную скорость снижения частоты встречаемости этого признака, исследователи, естественно, пришли к выводу, что объяснение быстрой эволюции березовых пядениц именно естественным отбором (под хищническим давлением птиц) является совершенно достаточным (Cook et al., 2012).
Действительно, компьютерное моделирование показывает, что при разнице в приспособленности 10%, частота встречаемости вредного признака за 40 лет должна упасть от исходных 90% до примерно 5%, если смена поколений осуществляется 1 раз в год.
Таким образом, озвучиваемые некоторыми верующими дарвинистами «десятки тысяч лет, необходимые для эволюции под действием естественного отбора» - это просто от незнания того предмета, который они пытаются защищать.
На самом деле, здесь всё зависит от степени преимущества в выживании, которое может обеспечить тот или иной признак.
Чтобы дальше не путаться, приведу примерный рейтинг ожидаемых скоростей естественного отбора в зависимости от степени биологического преимущества, которую даёт некий теоретический признак.
Итак:
1. Если некий (теоретический) признак даёт преимущество в выживании 100%. То есть, если особь, обладающая этим признаком, в среднем, оставляет вдвое больше потомства, чем особь, не обладающая этим признаком. Тогда практически полная замена в популяции особей, не обладающих этим признаком, на особей, обладающих этим признаком, должна произойти… всего за 20 поколений.[92] Что для живых существ, размножающихся один раз в год, составит всего 20 лет. А для организмов, размножающихся, например, три раза в год – вообще 7 лет.
Однако верующие дарвинисты скажут, что 100%-ное увеличение числа потомков – это, на самом деле, просто чудовищное биологическое преимущество, которое может обеспечить далеко не каждый признак. Поэтому, якобы, мы и не наблюдаем в природе таких молниеносных эволюционных переходов.
Тем не менее, даже такие признаки в природе – имеются. Например, переход от полового размножения к бесполому (или к партеногенезу) даёт именно немедленное двойное преимущество в числе потомков. Потому что в случае бесполого размножения или партеногенеза (или гермафродитизма) размножается каждая особь. На создание «бесполезных» (самостоятельно не размножающихся) самцов - ресурсы вообще не тратятся. Это обстоятельство и дает чудовищную (двойную) прибавку в скорости размножения таких организмов (Рис. 32):
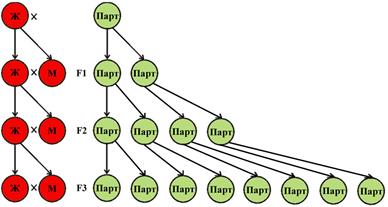
Рисунок 32. Увеличение численности партеногенетической популяции (зеленые кружки справа) в сравнении с популяцией, размножающейся «классическим» половым способом, с соотношением полов 50:50 (красные кружки слева). В данной модели каждая самка «классической» половой популяции (Ж) и каждая самка партеногенетической популяции (Парт) создаёт одинаковое число потомков, равное 2 (для наглядности). Но в «классической» половой популяции каждый второй потомок – это самец (М), который сам не разножается. В то время как в партеногенетической популяции размножаются все особи. В итоге, уже в третьем поколении (F3) число потомков первой партеногенетической самки составляет 8 особей, в то время как численность «классической» половой популяции – вообще не растет (при числе потомков каждой самки, равной двум). Очевидно, что при прочих равных, партеногенетические (или бесполые) популяции должны стремительно вытеснять «классические» половые популяции за счет огромной разницы в темпах размножения.
В связи с этим, в современной теоретической биологии существует так называемая «королева эволюционных проблем», где биологи пытаются ответить на вопрос, почему же столь чудовищно выгодное размножение, как бесполое, практически игнорируется видами, размножающимися половым способом.[93] Этой «королеве эволюционных проблем» посвящены десятки научных гипотез. Но ни одна из них пока не смогла ответить на заданный вопрос исчерпывающим образом. То есть, это очередной «эволюционный скандал» - когда в рамках теории эволюции должны побеждать организмы, оставляющие больше потомства… но реальные организмы, как говорится, «плюют с большой колокольни» на то, что они должны этой несчастной теории, и продолжают размножаться так, как им заблагорассудится.
Например, одна из гипотез причин существования полового размножения в природе – это необходимость постоянной борьбы с паразитами, в ходе «эволюционной гонки вооружений».[94] В рамках этой гипотезы предполагается, что половое размножение даёт более серьезную защиту от атак постоянно эволюционирующих паразитов или инфекций - за счет большего генетического разнообразия особей. В генетически разнообразных популяциях половых организмов обязательно найдутся отдельные особи, устойчивые к новому варианту инфекции. В то время как в генетически идентичных бесполых популяциях, новый успешный паразит может произвести страшное опустошение.
Но если это так, тогда бесполый (или партеногенетический) способ размножения мог бы особенно пригодиться высшим животным, например, позвоночным. Потому что у позвоночных имеется специфический способ борьбы с паразитами – чрезвычайно эффективная иммунная система, которая сама по себе создает разнообразие возможных антител для борьбы с инфекцией.
Непонятно почему, имея столь эффективный механизм борьбы с инфекциями, позвоночные животные так редко прибегают к вдвое более быстрому (или вдвое более экономичному) партеногенетическому размножению.
Более того. Помимо своей двойной мощности, партеногенетическое размножение еще и полностью избавляет животное от проблемы поиска полового партнера. А также от необходимости тратить время и энергию на какие бы то ни было «брачные игры». Соответственно, исчезает риск получения травм и инфекций при контакте с половыми партнерами (например, во время копуляции).
А ведь для позвоночных животных характерны крупные размеры тела, вследствие чего их темп размножения является низким (а в некоторых случаях экстремально низким). Понятно, что бесполое размножение или партеногенез (или хотя бы чередование бесполых и половых поколений) могло бы очень серьезно поправить эту проблему. Однако как раз среди позвоночных животных – партеногенез (или гермафродитизм) встречается экстремально редко (а бесполое размножение – вообще никогда).[95]
Честно говоря, весьма забавно читать такие гипотезы, где верующие дарвинисты пытаются объяснить сами себе, почему же такой чудовищно полезный признак, который даёт немедленное двойное преимущество (сразу +100% к приспособленности), тем не менее, не реализуется в популяциях организмов, размножающихся половым путем… И в то же время (те же самые) верующие дарвинисты убежденно рассказывают нам истории о том, как естественный отбор чутко замечает и кропотливо отбирает даже такие признаки, которые повышают приспособленность особи на 0.01% или даже на 0.001%. Например, появилось какое-нибудь дополнительное пятнышко на «шкуре» какого-нибудь очередного насекомого с защитной окраской, которое повышает его защиту еще чуть-чуть (на 0.01%). И верующий дарвинист убежденно говорит нам, что естественный отбор замечал такие пятнышки, и отбирал их. А потом еще и еще. И в результате, некоторые насекомые превратились примерно вот в такие шедевры маскировки (Рис. 33):

Рисунок 33. Очередной шедевр маскировки, который, согласно дарвиновской вере, появился путем «естественного отбора многих мелких изменений». Обратите внимание на ряд темных пятен на надкрыльях этого кузнечика, которые изумительно имитируют гниение листа растения.
И подобные увлекательные сказки дарвинисты рассказывают нам, в то время как реальные факты говорят о том, что естественный отбор может в упор не замечать даже такие возможности, которые повышают приспособленность особей не то что на гипотетические 0.01%, но даже на совершенно реальные 100% (что в 10000 раз больше). Причем такое преимущество в случае перехода к бесполому размножению реализовалось бы немедленно. Но что-то не торопится естественный отбор исполнять свои прямые обязанности даже в этом случае. Что уж тогда говорить о гипотетических 0.01% преимущества.
Вообще, с помощью «королевы эволюционных проблем» можно довольно легко опровергнуть всю теорию естественного отбора (или хотя бы поставить её под очень большое сомнение), несмотря на большое число гипотез, пытающихся «вытащить» дарвинизм из этого «эволюционного скандала». Для этого достаточно вспомнить не о чисто бесполом размножении, а о возможности чередования полового и бесполого поколений (метагенез) или о возможности чередования полового и партеногенетического поколений:
1. Такой способ размножения сохраняет все предполагаемые плюсы полового размножения (которые предполагаются в сооветствующих гипотезах). Но при этом темпы размножения всё еще очень высоки. Даже при постоянном чередовании полового поколения с бесполым (или с партеногенетическим), темп размножения таких видов всё еще будет в 1.5 раза выше, чем у видов, размножающихся только половым способом. Потому что фактическое соотношение размножающихся особей (к неразмножающимся самцам) – будет 2:1 (одна половая и одна бесполая или партеногенетическая «самка» на одного самца), вместо 1:1 в «классическом» половом размножении.
2. При этом поколение бесполых (или партеногенетических) «самок» будет полностью «иммунно» к «засорению» своих рядов бесполезными самцами. Что делает известный «принцип Фишера», выдвинутый в оправдание соотношения полов 50:50 (которое наблюдается в природе для подавляющего большинства видов) - совершенно непригодным в случае чередования полового и партеногенетического поколений.
«Принцип Фишера» состоит в том, что если в популяции преобладают самки, то в такой популяции становится выгодно быть самцом, потому что в этом случае самец имеет возможность передать следующему поколению гораздо больше своих генов, чем среднестатистическая самка. Поэтому в таких популяциях доля самцов в ходе эволюции должна постепенно повышаться. Вплоть до того момента, пока соотношение полов не станет 50:50. Потому что при более высоком соотношении самцов, быть самцом становится уже не выгодно – среднестатистическая самка в любом случае оставит своё потомство, в то время как не каждый среднестатический самец сможет оставить свое (из-за конкуренции между самцами). Вот поэтому, согласно «принципу Фишера», соотношение полов у подавляющего числа видов оказывается равным (50:50), несмотря на то, что для популяции в целом такое соотношение полов не является выгодным. Ведь чем выше в популяции доля самок, тем быстрее такая популяция способна наращивать численность.
Однако при чередовании поколений, соотношение «полов» в популяции следующее: 1 бесполая «самка» : 1 половая cамка : 1 cамец. Такое соотношение полов будет эволюционно стабильным, так как самцы не могут «проникнуть» в бесполую (или партеногенетическую) часть популяции (и нарастить там свою долю). Что полностью снимает аргументацию «принципа Фишера». Тем не менее, темпы размножения такой популяции всё еще будут в 1.5 раза выше темпов «чистого» полового размножения.
3. Более того, если половые самцы и самки будут являться гаплоидными, то на этой (гаплоидной) стадии мог бы происходить эффективный очищающий отбор, что позволило бы таким популяциям вообще не накапливать груз вредных мутаций, избавляясь от таких мутаций сразу же, по мере их появления.
А для эффективного избавления от слабо-вредных мутаций в популяциях таких видов были бы задействованы точно такие же механизмы, которые существуют в любой половой популяции, теоретически считаясь «преимуществами полового размножения».[96] Единственным отличием здесь будет то, что эти механизмы в данном случае будут осуществляться не в каждом поколении, а через одно.
И вот такой способ существования вида обеспечил бы темп его размножения в 1.5 раза выше скорости размножения любого «классического» полового вида.
Тем не менее, чередование половых и бесполых поколений в природе – это весьма экзотический способ размножения, который практикуется сравнительно малым числом биологических таксонов. В то время как в рамках дарвинизма, размножение путем чередования половых и бесполых (или партеногенетических) особей, по идее, должно безраздельно господствовать в живой природе (над просто половым размножением). По этому поводу можно даже переформулировать «королеву эволюционных проблем» соответствующим образом – почему в природе настолько мало распространено не только бесполое размножение или партеногенез, но даже чередование полового поколения с бесполым (или партеногенетическим)?
Хотя, казалось бы, эволюционное возникновение описанной схемы размножения является не слишком сложной задачей. Например, среди позвоночных животных существует несколько десятков партеногенетических видов (примерно 0.1% от общего числа всех видов позвоночных животных). Партеногенез установлен у отдельных видов рыб, амфибий, рептилий и даже птиц. Некоторые виды ящериц вообще размножаются только партеногенетически. У других видов позвоночных, несмотря на преобладание партеногенетического размножения, тем не менее, регулярно появляются и самцы, и осуществляется половой способ размножения. А у третьих видов (партеногенетических) позвоночных животных – наоборот, преобладает обычный половой процесс, но партеногенез, тем не менее, наблюдается или эпизодически, или в случае крайней необходимости (например, при отсутствии половых партнеров).
Казалось бы, эти факты указывают на то, что никаких особенных препятствий для эволюционного перехода к партеногенетическому размножению, или к размножению со сменой партеногенетического и обычного полового поколений – не имеется. И мы должны были бы наблюдать такие виды в изобилии. Однако мы наблюдаем то, что наблюдаем – крайнюю редкость подобных форм размножения в природе, несмотря на их (теоретически) огромную эффективность в сравнении с обычным половым размножением.
Впрочем, эту тему следует подробно рассмотреть где-нибудь в другом месте. Сейчас же мы пока просто описываем возможные скорости (воображаемой) эволюции в зависимости от силы естественного отбора.
Второй пример, когда особь могла бы обеспечить себе двойное преимущество в размножении (или даже тройное и четверное) – это знаменитые лососи. Как известно, почти все виды тихоокеанских лососей (род Oncorhynchus) запрограммированно погибают сразу же после первого размножения. Например, такие представители этого рода, как горбуша, кета, кижуч, нерка и чавыча[97] массово гибнут после первого же нереста в период от нескольких дней до двух-трех недель. Этим данные виды лососей резко отличаются от своего близкого родственника и соседа – от стальноголового лосося (Oncorhynchus mykiss), который после первого размножения массово помирать даже и не думает, а продолжает жить и размножаться (в следующие сезоны размножения).
Такое странное, массовое и явно генетически запрограммированное «харакири» целого ряда видов тихоокеанских лососей, конечно, не может не вызывать удивление. А если говорить прямо – вообще опровергает эволюцию по сценарию преимущественного выживания более приспособленных. Потому что в рамках теории, где преимущественно выживают более приспособленные особи, не может закрепиться такой признак, который приводит к запрограммированной гибели этих особей. В рамках дарвинизма, легко можно представить себе вынужденную смерть организма, от каких угодно естественных причин (например, от истощения после размножения). Но нельзя представить в рамках этой теории возникновение специальной генетической программы, которая в какой-то момент включается, и гарантированно убивает организм. Ибо невозможно представить признак, более вредный для организма, чем запрограммированная смерть этого организма. Поэтому наличие специальных генетических программ, приводящих к гарантированной смерти, нельзя объяснить в рамках теории выживания более приспособленных. Тем не менее, массовая гибель тихоокеанских лососей выглядит именно так, как будто эта гибель генетически запрограммирована.
В этом смысле интересно, что еще один вид тихоокеанских лососей – сима (Oncorhynchus masou) тоже массово погибает сразу после первого же размножения. То есть, в этом отношении данный вид почти копирует других своих тихоокеанских коллег. Но в отличие от полных «камикадзе», особи симы погибают всё-таки не все. Некоторые особи благополучно переживают первое размножение. И в этом случае, такие особи потом участвуют в размножении второй раз.
Но ведь если некая особь не погибнет (запрограммированно) сразу после первого размножения, а размножится еще и второй раз, то это значит, что она оставит после себя ровно вдвое больше потомков, чем её «одноразовые» конкуренты. Это и есть то самое 100%-ное преимущество, которое получит такая («двуразовая») особь. Если этот признак окажется наследуемым, то это должно привести к тому, что сима (O. masou) из «одноразового» вида превратится в «двуразовый» вид всего за 20 поколений! Что конкретно для симы равняется примерно 60-80 годам. То есть, мы могли бы (в принципе) наблюдать буквально молниеносные эволюционные изменения биологических видов… если бы теория эволюции по дарвиновскому механизму была правдой, а не чисто теоретической спекуляцией.
Даже если предположить, что до второго цикла размножения будет доживать всего 10% взрослых особей (погибая от разных причин в течение года), то и тогда получается, что «двуразовость» лучше «одноразовости» на 100/10=10%. В этом случае практически полная замена «одноразовых» особей «двуразовыми» должна наблюдаться примерно за 100–150 поколений. Так почему же тихоокеанские лососи до сих пор запрограммировано дохнут в массовом порядке?
Примерно таким же чудаковатым образом ведет себя и семга (атлантический лосось, Salmo salar). В отличие от подавляющего большинства своих тихоокеанских коллег, семга не погибает (запрограммировано) сразу после первого нереста, а может размножаться неоднократно (так же как и уже упомянутый выше стальноголовый лосось). Однако лишь незначительная часть семги доживает до второго размножения. Потому что организм далеко не каждой семги выдерживает те нагрузки, которые семга добровольно возложила на себя в ходе собственного размножения. А именно, семга, зайдя в реку, полностью перестаёт питаться. Поэтому она вынуждена осуществлять весь нерест на своих «внутренних резервах». В результате, значительная часть особей истощается настолько, что гибнет сразу после первого нереста. Особенно в этом отношении достаётся той семге, которая заходит для размножения в реку очень рано, поэтому вынуждена торчать в этой реке (не питаясь!) чуть ли ни целый год, дожидаясь периода нереста, приходящегося на осень. Зачем семга при этом добровольно отказывается от еды, известно только самой семге.
И вот если истощенная рыба всё-таки не сдохнет сразу после нереста, а дотянет в живом виде обратно до моря, то там к ней снова приходит аппетит. Семга быстро поправляется, и через год может направиться на следующий нерест.
Интересно, насколько бы повысилась приспособленность семги, если бы какая-нибудь мутантная особь наконец «догадалась», что и в реке тоже можно питаться. Тем более что сама семга в молодости занимается именно этим делом (в этой же самой реке на протяжении 1-3 лет). И тем более, что в природе существуют многочисленные пресноводные популяции семги, которые живут в пресных озерах всю свою «сознательную жизнь», и в этой пресной воде не теряют аппетита.
Понятно, что если бы какая-нибудь семга начала питаться прямо в процессе своего «великого похода к нерестилищу», то это бы повысило приспособленность семги достаточно заметным образом, чтобы такие особи стали вытеснять не питающихся особей в ходе естественного отбора. И в результате, этот биологический вид мог бы эволюционно измениться. Именно так и должно быть в том воображаемом фантастическом мире, в котором эволюция более приспособленных, действительно, существует. Но мы с Вами живем в реальном мире, где семга остаётся семгой.
Вообще, в отношении «одноразового размножения», нашей любимой эволюции можно было бы широко развернуться при желании. Ведь такими «одноразовыми» являются далеко не только лососи. А еще и многие головоногие моллюски. Считается, что кальмары, каракатицы и осьминоги (за исключением аргонавтов) – тоже все «одноразовые» (Несис, 2005:135). Во всяком случае, подавляющая их часть.
Это просто удивительная расточительность. Ведь головоногие моллюски являются одними из самых высокоорганизованных живых существ. А некоторые еще и одними из самых высоко интеллектуальных животных. Головоногих моллюсков часто так и называют – «приматы моря». И вот представьте себе, что этот «примат моря» рос-рос, рос-рос, и вырос, например, в тихоокеанского осьминога, размером около 2 метров и массой около 30 кг (!) для того, чтобы в первый раз размножиться… и вскоре после этого умереть!
Это всё равно, как если бы человек копил на дорогую машину в течение 10 лет, наконец, накопил бы на неё, приобрел в дорогом салоне, потом покатался бы один день (!) и после этого… сразу сдал в утиль.
Однако в отношении человека такое поведение всё же возможно. Может быть, этот человек просто большой чудак. Но как понять такой удивительный признак в рамках теории естественного отбора? Казалось бы, если уж ты сумел пережить всех своих многочисленных братиков и сестер, погибших на самых разных стадиях взросления (в основном, на стадии самой маленькой мелюзги), то теперь ты (достигший почти вершины пищевой пирамиды), просто обязан размножаться «до упора», пока смерть по независящим от тебя обстоятельствам не прекратит этот процесс размножения. То есть, если уж сумел вырасти таким большим и сильным, то в рамках дарвинизма, должен теперь заниматься копированием собственных «эгоистичных генов» так долго и так много, насколько это вообще возможно. Однако нет. Лососи, осьминоги, кальмары и каракатицы, видимо, просто не слышали о дарвинизме. Поэтому запрограммировано дохнут сразу после первого размножения.
В растительном мире таким же порочащим дарвинизм образом жизни занимается широко известный бамбук. Он может долго расти и размножаться вегетативным способом. И в результате сильно разрастается. Но стоит ему только зацвести (один раз в несколько десятков лет!), как все надземные части этого растения «дохнут» сразу после этого (первого) размножения. А поскольку бамбук имеет привычку зацветать очень дружно, то таким образом можно за один раз лишиться сразу целой бамбуковой рощи. И это весьма удивительно в рамках концепции биологической выгоды. Ведь получается, что после своего первого и единственного полового размножения, бамбуковые рощи… дружно освобождают все территории, которые они успешно захватили (победив всех конкурентов). И теперь лишь воле случая предстоит решить дальнейшую судьбу этих территорий. Причем не факт, что в жестокой конкуренции здесь снова победят потомки именно исходных растений (которые здесь росли раньше), а не какие-то другие. Отсюда можно сделать вывод, что бамбук (род растений, включающий более 100 видов) появился как угодно, но только не при участии пресловутых «эгоистичных генов» Докинза. Ибо с позиций «эгоистичных генов», уже захваченную территорию (с ресурсами, позволяющими плодить и плодить новые копии «эгоистичных генов») - добровольно терять такую территорию (начиная всё сначала) - просто верх безумия.[98]
И всё же самым большим оригиналом в этом отношении является недавно открытая мадагаскарская пальма Tahina spectabilis. Это чудо анти-дарвиновской «эволюции» является одной из крупнейших пальм Мадагаскара, достигая высоты 18 метров. Предполагается, что она достигает такой высоты примерно за 50 лет. И вот после того, как все эти предполагаемые 50 лет пальма росла (видимо, конкурируя с другими растениями?) и наконец, ей удалось вырасти в большое красивое дерево… После этого она отнюдь не размножается каждый год, давая семена в течение последующих 50 лет (как, например, кокосовая пальма). Нет, для неё это, видимо, скучно. Вместо этого данная пальма сразу производит столько цветов… что погибает в течение нескольких недель после первого же плодоношения.
Что и говорить, это очень интересное поведение для пальмы.
Но чтобы с этим разобраться до конца, пожалуй, лучше начать с самого начала.
Самое начало началось в уже далеком 1954 году, когда биолог Л.С. Коул опубликовал работу, результаты которой произвели большое впечатление на биологов того времени. В этой работе Коул задался целью ответить на вопрос, что выгодней - размножаться только один раз в жизни (а потом помереть), или же размножаться много раз. В результате сравнительно несложных математических расчетов, Коул пришел к выводу, что размножаться всего один раз в жизни столь же выгодно (для некоей линии организмов), что и размножаться много раз – если «одноразовый» организм будет производить (за свой единственный акт размножения) всего на одного (!) потомка больше, чем «многоразовый» организм. Отсюда следует, что если «одноразовый» организм произведёт (за раз) не на одного потомка больше, а хотя бы на двух (не говоря уже о трех или о десяти), то размножаться единственный раз в жизни становится выгодней, чем быть «многоразовым» организмом.
Например, у нас имеется некий «многоразовый» биологический вид, который производит за один раз вообще минимальное число детей – всего одного потомка за один сезон размножения. Тогда «одноразовому» организму для того чтобы догнать «многоразовый» вид по темпам размножения (темпам прироста численности), необходимо «сделать» за один раз не менее 2 потомков. Но два потомка вместо одного (за сезон размножения) – это двойное репродуктивное усилие (усилие, потраченное на размножение) в сравнении с «многоразовым» видом. А это очень серьезная разница. Далеко не каждый организм, наверное, на это способен (см. ниже). Поэтому здесь преимущество «одноразового» вида над «многоразовым» отнюдь не кажется очевидным.
Но стоит нам взять какой-нибудь другой пример, как ситуация сразу же парадоксально меняется. Дело в том, что неважно, сколько потомков мы возьмем – одного потомка, или десять, или сотню, или тысячу, или даже сто тысяч (каких-нибудь икринок). Но достаточно добавить к этому числу всё того же (единственного!) дополнительного потомка… как темп размножения «одноразового» и «многоразового» организма тут же выравнивается!
То есть, по темпам размножения виды получаются совершенно равными, если за один сезон они производят следующее число потомков:
1). Многоразовый вид 1 потомка, а одноразовый вид – 2 потомков.
2). Многоразовый вид 10 потомков, а одноразовый вид – 11 потомков.
3). Многоразовый вид 100 потомков, а одноразовый вид – 101 потомка.
4). Многоразовый вид 1000 потомков, а одноразовый вид – 1001 потомка.
5). Многоразовый вид 10000 потомков, а одноразовый вид – 10001 потомка.
6). Многоразовый вид 100000 потомков, а одноразовый вид – 100001 потомка.
И так далее.
Но что такое прибавка, например, к исходной тысяче икринок – еще одной икринки (тысяче первой)? Это ведь совершенно ничтожное дополнительное репродуктивное усилие, которое отличается от производства исходных 1000 икринок всего на 0.1%.
Поэтому «одноразовый» вид, казалось бы, получает очень серьезное преимущество. Ведь он не обязан выживать после своего единственного сезона размножения. «Одноразовая» особь может спокойно «пойти вразнос», бросив вообще все ресурсы на создание потомства. И в результате, «одноразовая» особь окажется способна выделить на размножение, возможно, на 30% больше ресурсов, чем «многоразовая» особь? В то время как «многоразовая» особь должна беречь ресурсы, чтобы выживать и дальше, после сезона размножения. Поэтому «многоразовые» особи по определению должны больше экономить на ресурсах, выделяемых на размножение, чем «одноразовые» виды. Но в случае дополнительных 30% ресурсов, брошенных на размножение, у «одноразовой» особи получится уже примерно 1300 икринок вместо 1000 икринок, которую может позволить себе «многоразовая» особь. Насколько же это выгодней, если уже 1001 икринка – уравнивает темпы размножения этих видов?
Этот математический вывод насчет «плюс всего одного потомка» (которого уже достаточно, чтобы полностью уравнять темпы размножения «одноразовых» и «многоразовых» видов), показался биологам парадоксальным. Ведь интуитивно кажется, что тот, кто размножается много раз (каждый раз порциями по 1000 икринок), должен увеличивать свою численность больше, чем тот, кто размножается только один раз, производя при этом 1001 икринку. Тем не менее, математически, расчеты Коула были верны.
Те, кому эти расчеты всё-таки не кажутся верными, могут легко проверить их при наличии хотя бы простейших навыков программирования. Соответствующие алгоритмы, которые будут показывать изменение числа особей в разных поколениях в случае «многоразовых» и «одноразовых» организмов – очень простые, и могут быть легко выполнены на компьютере. Ну а те, кому это делать лень, могут поступить еще проще. Просто представьте себе такую ситуацию, когда размножение «многоразового» организма начинается всего с одной единственной особи. В этом случае легко понять, что эта единственная особь, оставшаяся после первого сезона размножения, и приступившая к своему второму размножению в следующем сезоне, по сути, будет выступать в роли того самого «единственного дополнительного потомка», который необходим «одноразовому» организму для выравнивания его темпов размножения с «многоразовым» видом.
Итак, расчеты Коула верны. Но тогда получается, что быть одноразовым – гораздо выгодней, чем быть многоразовым. И это тоже выглядит парадоксальным, потому что тогда возникает вопрос – почему же все организмы, которых мы наблюдаем в природе, не практикуют одноразовое размножение, с максимальным репродуктивным усилием в ходе этого размножения? Почему в природе так много организмов, размножающихся неоднократно?
Именно вследствие своей парадоксальности, выводы Коула получили название «парадокса Коула», и под таким названием вошли в историю биологии.
Однако сегодня «парадокс Коула» - это уже и есть только история биологии. Потому что в более поздних работах (Charnov & Schaffer, 1973; Yang, 1981) этот парадокс был разрешен. Авторы этих работ показали, что Коул в своих расчетах не учел, прежде всего, разницу между выживаемостью взрослых особей и выживаемостью потомства. А это очень важное обстоятельство.
Расчеты Коула сохраняют справедливость только при одинаковой выживаемости взрослых особей и потомства. Если же выживаемость взрослых особей существенно выше выживаемости потомства, тогда преимущество получают «многоразовые» особи, и преимущество тем более значительное, чем сильнее эта разница (в выживаемости взрослых и детей). Действительно, если родительская особь (оставшаяся от предыдущего поколения) ничем не отличается по эффективности выживания от своих детей, то эту родительскую особь в следующие периоды размножения можно спокойно рассматривать просто в качестве еще одного (дополнительного) потомка. Если же эта взрослая особь, допустим, имеет в тысячу раз большие шансы дожить до следующего сезона размножения, чем каждый из его детей (взятый в отдельности), то разве тогда можно оперировать (в соответствующих расчетах) этой особью наравне с её детьми? Очевидно, что в этом случае взрослая особь в тысячу раз ценнее каждого из своих детей. И именно такой её «вес» должен учитываться в расчетах.
Что может увеличивать шансы отдельной особи успешно выживать?
Во-первых, этому очень способствует большой размер организма. Например, каких-нибудь полевых мышей едят все кому не лень. Поэтому шансы каждой отдельной мыши прожить свою жизнь долго и счастливо - весьма призрачны (во всяком случае, в отношении слова «долго»). А вот, например, африканский лев уже вполне может рассчитывать на целый ряд лет успешной взрослой жизни.
Во-вторых, на шанс долгой жизни весьма положительно влияет сложность организации конкретного живого существа. Чем сложнее устроено существо, тем эффективней оно избегает разных неблагоприятных ситуаций. Например, хорошо развитые органы чувств помогают и эффективно избегать хищников, и самому эффективно питаться (не становясь жертвой голода).
Таким образом, здесь можно сформулировать некое общее правило – чем больше по размеру, и чем сложнее устроен живой организм, тем выше эффективность выживания каждой отдельной особи этого вида. Следовательно, тем выше ценность этой (взрослой) особи для размножения. И, следовательно, тем выгодней такому организму использовать «многоразовую» стратегию размножения.
И наоборот, чем меньше организм и чем проще он устроен, тем меньше «стоимость жизни» каждой отдельной особи. И тем меньше эта стоимость отличается от стоимости жизни его детей. Следовательно, такой организм мог бы приобрести выгоды от «одноразовой» стратегии размножения.
Таким образом, такие сложно устроенные существа, как головоногие моллюски или лососи – вряд ли поступают целесообразно, запрограммировано подыхая сразу после первого размножения.
И уж тем более странно поступает в этом отношении мадагаскарская «пальма-самоубийца», которая имеет настолько большой размер, что достигает этого размера только к 30-50 годам. Но при этом размножается только один раз.
Второе обстоятельство, которое не учел в своих расчетах Коул – это специфику условий природной среды. Например, если условия среды такие, что чреваты внезапной смертью (от каких-нибудь «причин неодолимой силы»), то такая обстановка не способствует «многоразовости». В таких условиях было бы здорово дожить хотя бы до первого размножения (как можно быстрее), и вложиться в него максимально, потому что до следующего размножения можно уже не дожить.
И наоборот, если окружающие условия благоприятствуют именно взрослым стадиям организмов, и не благоприятствуют детским стадиям, тогда, безусловно, будет выгодней многоразовое размножение. Например, в каком-нибудь тропическом лесу, взрослое дерево, уже сумевшее захватить все необходимые ему ресурсы, невообразимо ценнее каждого конкретного семечка, производимого им. Потому что шансы конкретно этого семечка превратиться во взрослое дерево, захватив (кем-то оставленное) пространство в этом тропическом лесу – стремятся к нулю.
Поэтому мадагарская пальма-самоубийца, погибающая после своего первого же размножения (на радость всем своим конкурентам) в условиях тропиков, выглядит в высшей степени странно (в рамках представлений о биологической целесообразности).
Количественно всё выше сказанное можно легко оценить, если принять следующее допущение – если мы допустим, что численность особей тех или иных биологических видов в природе остаётся примерно постоянной на протяжении длительных периодов времени.
Скорее всего, так и есть. Ведь мы не наблюдаем зарастания нашей Земли, например, кокосовыми пальмами? Или полного забивания всех дальневосточных рек триллионами лососей? Но если это так (если численность озвученных видов в природе остаётся примерно постоянной), тогда «вес» взрослых особей этих видов для размножения оценить достаточно легко.
Например, какая-нибудь самка чавычи отметала свои 10.000 икринок, и после этого с чувством выполненного долга… померла. Поскольку мы приняли, что численность чавычи в природе остаётся примерно постоянной, то отсюда следует, что из этих 10.000 икринок, отложенных самкой чавычи в этом году, в конечном итоге (через несколько лет) доживет до размножения (в среднем)… только одна самка.
Следовательно, вероятность доживания до размножения каждой отдельной икринки в данном случае составляет всего 0.01%. Это ничтожно малая величина. Поэтому приравнивать ценность такой икринки к ценности огромной взрослой особи чавычи (весом до 50 кг), находящейся почти на вершине пищевой пирамиды – ни в коем случае нельзя. А можно поступить просто – взять все эти 10.000 икринок, и просто приравнять их всех вместе к «стоимости» одной взрослой самки.
Тогда у нас получится, что самка чавычи порождает (условно говоря) просто следующую самку чавычи (в виде «суммы» из 10.000 икринок). И после этого погибает.
Теперь давайте представим себе гипотетическую мутантную чавычу, которая не погибает после первого размножения, а размножается второй раз на следующий год, отметав те же самые 10.000 икринок. Очевидно, что такая самка будет являться более приспособленной, чем «одноразовая», нормальная самка. Чтобы «одноразовая» самка выровнялась с этой мутантной «многоразовой» самкой в темпах размножения, она должна выметать за один раз не 10.000 икринок, а 20.000 икринок, что будет означать («в переводе» этих икринок на взрослых самок) – 2 самки. Но сможет ли самка чавычи выносить и выметать в два раза больше икры, чем многоразовая особь?
Ведь «многоразовые» виды рыб (которых вообще-то большинство) тоже, наверное, напрягаются, стремясь отметать как можно больше икры. Так сумеют ли «одноразовые» особи создать вдвое больше икры, чем многоразовые, чтобы уравнять свои шансы?
Это трудно оценить напрямую. Но приблизительно оценить все-таки можно.
Воспользуемся для этого спортивной штангой. Допустим, я поднимаю от груди спортивную штангу с максимальным весом 160 кг (всего один раз). Поднять больше 160 кг, или поднять 160 кг больше одного раза – я уже не могу. Однако при этом я могу поднять 145 кг… примерно 4-5 раз! То есть, прямая арифметика не действует в случае живых организмов. Если я поднимаю 160 кг только один раз, то из этого совсем не следует, что я смогу поднять 80 кг всего 2 раза. Нет, 80 кг для меня – это очень легкий разминочный вес, который мне надо поднимать и поднимать (примерно раз 30), чтобы устать. Более того, я могу поднять 145 кг 4-5 раз, хотя 145 кг меньше 160 кг… всего примерно на 10%!
Что же получается, я могу без особенного напряжения поднимать вес, который меньше моего максимального веса… всего на 10%. И завтра приду, и опять подниму этот вес (тоже без особого напряжения). И еще послезавтра повторю то же самое.
Более того, сегодня я могу поднять даже свой самый тяжелый вес – 160 кг (один раз). Мне будет очень тяжело, но при этом я всё-таки не умру от напряжения. А раз так, то потом я смогу отдохнуть, и через неделю подниму этот же вес снова.
Более того, если я – еще молодой юноша, то через пару лет я могу еще и подрасти, и тогда я подниму уже не 160 лет, а даже больше. Так стоит ли мне прямо сейчас надрываться так, чтобы умереть?
И могу ли я утверждать, что если бы я всё-таки решил, что именно сегодня «есть мой последний и решительный бой», то от осознания особенной важности этого дня, я бы вдруг поднял не 160 кг, а сразу 180? Я глубоко сомневаюсь в возможности такого события. Скорее всего, штанга весом 180 кг придавила бы меня к спортивной скамье, как таракана. Несмотря на то, что 180 кг – это всего на 12% больше того предельного веса, который я вполне способен выжать от груди один раз.
Может быть, примерно такие же зависимости существуют и в отношении репродуктивных усилий?
Тогда «многоразовые» особи, достаточно напрягшись, смогут произвести 10.000 икринок. А «одноразовые» особи, совершив даже смертельное усилие, не смогут произвести икринок больше чем на 10-20% от того количества, которое произвели их «многоразовые» коллеги. То есть, не более 11000-12000 икринок.
Но тогда, в «переводе на взрослых самок», такое количество икринок составит всего лишь 1.1-1.2 самки. Но никак не 2 самки, которые необходимы для того, чтобы «одноразовость» тихоокеанских лососей стала биологически оправданной.
Однако не будем торопиться с выводами. Ведь в наших расчетах мы пока еще не учитывали смертность взрослых особей. Пока они у нас были как бы бессмертными, то есть, взрослые особи у нас пока доживали до следующего сезона размножения с вероятностью 100%. Что не очень реалистично. Поэтому давайте введем определенный шанс доживания взрослой особи до следующего периода размножения.
И вот если мы постановим, что этот шанс достаточно низок, например, на уровне 10-20% (то есть, только одна из пяти или даже одна из десяти взрослых особей доживает до следующего размножения), тогда темпы размножения «одноразовых» и «многоразовых» особей опять сравняются. То есть (уже задерганная нами) самка «одноразовой» чавычи, совершившая сверхмощное репродуктивное усилие, и в результате выметавшая 11000-12000 икринок за своё первое (и последнее) размножение… и воображаемая «многоразовая» чавыча, выметавшая 10000 икринок и после этого оставшаяся жить… эти особи будут иметь примерно одинаковую приспособленность, если вероятность «многоразовой» чавычи дожить до следующего сезона размножения примерно 10-20%.
Так это или не так на самом деле, то есть, эквивалентны ли реальные цифры ежегодной смертности взрослых лососей – воображаемому проценту дополнительных икринок – пусть на этот вопрос отвечают специалисты. Нам это не слишком важно. Нам важно другое. Мы сейчас оперировали рассуждениями про «доступные ресурсы» и «репродуктивные усилия», игнорируя то важное обстоятельство, с которого, собственно, начали. Ведь лососи гибнут не от истощения (или «надорвавшись от усилий»). Складывается такое ощущение, что лососи специально «хотят» умереть. И прилагают для этого все усилия.
Например, мы только что сделали вывод, что «одноразовость» обязательно предполагает дополнительный вклад ресурсов в размножение, по сравнению с «многоразовостью». Иначе «одноразовость» будет невыгодной. Но с этой точки зрения, твердое «решение» лососей не питаться (ни в коем случае!) во время собственного путешествия к нерестилищу – выглядит в высшей степени странным. Каковы бы ни были шансы конкретной рыбы, идущей на нерест, перекусить местным представителем речной фауны, зачем отказываться от такой возможности специально? Неужели категорический отказ от пищи помогает выметать больше икринок?
Более того, непосредственно перед нерестом, тела тихоокеанских лососей переживают серьезное перерождение (особенно у самцов). Челюсти искривляются, зубы увеличиваются (зачем?). Тело становится более высоким. У отдельных видов вырастает большой горб. Резко изменяется окраска – у одних видов тело становится очень ярким (ярко-красным или даже лилово-красным), а у других видов окрашивается в интенсивные темные цвета. Зачем такое перерождение нужно? Неужели оно как-то помогает увеличить число отложенных икринок?
А после нереста ткани лососей быстро деградируют. Такое ощущение, что лососи после нереста стремительно стареют. И гибнут именно по этой причине, а не в результате истощения. Причем смерть настигает их везде. Не важно, отнерестились ли они в самых верховьях какой-нибудь длинной реки, преодолев для этого целую кучу порогов. Или же в низовьях маленькой и спокойной речки, пройдя вверх (по спокойной воде) всего километров двадцать. Всё равно после нереста погибают все особи.
В общем, всё выглядит так, как будто это генетически запрограммированный процесс – специальное 100%-ное убийство взрослых особей сразу после первого размножения, не имеющее других целей, кроме самого этого убийства.
Вот об этой (специальной) генетической программе «гарантированного самоубийства» и идет сейчас речь. Такая программа не могла возникнуть под действием естественного отбора. Не могла просто потому, что гарантированно убивая особь, эта программа вообще не дает ни одной (даже самой удачливой) особи никаких шансов дожить до следующего размножения. Хотя если бы такая программа просто не работала, то особь могла бы выжить, и дожить до следующего размножения. И даже если бы такой шанс был всего 1%, то это уже всё равно было бы выгодно.
Тем не менее, тихоокеанские лососи продолжают дружно умирать.
Как такой признак мог появиться в ходе эволюции по механизму «преимущественного выживания более приспособленных» - совершенно непонятно.
И чтобы совсем уже закончить с темой «одноразовых лососей», отмечу еще одну «народную версию». В народе бродит популярная гипотеза, что лососи запрограммированно гибнут сразу после нереста якобы потому, что «лосось удобряет речку своей мертвечиной, это приводит к росту микроорганизмов, которыми кормятся его мальки» (я привел прямую цитату с одного из интернет-форумов).
Весьма высока вероятность, что и Вам тоже кто-нибудь такое заявит (если Вы, конечно, решите обсудить с верующим дарвинистом лососиную тему). Если такое случится, просто озвучьте своему дарвинисту следующий текст:
Эта версия, хотя и популярна в народных массах, но совершенно не выдерживает критики по следующим причинам:
1). Мальки лососей вылезают из-под камней (переходя к свободному питанию) лишь спустя 5-10 месяцев после того, как сдохнут их родители. А поскольку реки, где нерестится лосось, имеют, как правило, хорошее течение, то это значит, что сдыхающие родители кормят (своими разлагающимися телами) кого угодно, но только не собственных детей.
2). Даже если бы лососи действительно кормили собственных детей таким экстравагантным способом, то это бы все равно никак не помешало соответствующей эволюции. Если среди «одноразовых» лососей появится мутантный «многоразовый» лосось, то он получит в этом случае даже большую биологическую выгоду. Потому что лично его дети - тоже будут пользоваться этой «удобренной рекой» - наравне со всеми остальными мальками. В то время как взрослая (мутантная) особь сохранит себя до следующего размножения. Такая эгоистичная стратегия в данном случае - крайне выгодна, и поэтому должна моментально захватывать популяции лосося.
3). В этом случае непонятно, что «удобряют» (своими телами) кальмары, каракатицы и осьминоги, которые являются столь же «одноразовыми», как и лососи, но живут в океане. Неужели они пытаются удобрить мировой океан?!
4). Наконец, имеются озера, в которых всё рыбье население представлено исключительно щуками. Где взрослые щуки даже и не думают «удобрять» своими телами это озеро, подкармливая собственных (и чужих) детей, а вместо этого, наоборот, с удовольствием жрут любых щучек более мелкого размера. И так и живут. Тем не менее, щуки в этих озерах - не то что не вымирают, а наоборот, похоже, вытеснили вообще всех других рыб. Возникает вопрос - почему же щуки прекрасно обходятся без «удобрения» собственными телами этих озер, а лососи должны непременно удобрять реки (с быстрым течением)?
Я надеюсь, после прочтения этих четырех пунктов у Вашего дарвиниста больше не появится желание утверждать, что «лососям выгодно запрограммированно погибать, потому что лососи тем самым удобряют реки».
Ну а теперь давайте прикинем наши расчеты еще и в отношении уже упомянутой нами «пальмы-самоубийцы». Давайте сделаем это, просто сравнив их с известными всем кокосовыми пальмами. Пусть кокосовая пальма каждый год производит всего по 20 кокосовых орехов в течение 50 лет.
Эти 50 лет успешного плодоношения кокосовой пальмы можно, в принципе, представить, как существование с вероятностью умереть 2% в год.
В итоге, кокосовая пальма произведет за всю жизнь, в среднем, 1000 орехов. Поскольку мы не наблюдаем увеличения кокосовых пальм в природе, то это значит, что из 1000 орехов, в успешную взрослую пальму сумеет превратиться, в среднем, всего лишь одна кокосовая пальма. То есть, шанс превращения конкретного кокосового ореха во взрослую пальму в нашем случае примерно 0.1%. Таким образом, одна взрослая кокосовая пальма эквивалентна тысяче кокосовых орехов.
А раз так, то для того, чтобы сравняться с многоразовой кокосовой пальмой, гипотетической одноразовой пальме нужно за один раз произвести столько кокосов, чтобы это было эквивалентно хотя бы одной взрослой особи. То есть, вырастить за один раз на себе сразу 1000 полновесных кокосов. Понятно, что никакая, даже самая «одноразовая» пальма с такой задачей не справится. Поэтому мадагаскарская пальма-самоубийца, погибающая сразу после первого и единственного в её (долгой) жизни размножения, гарантированно имеет во много раз более низкие темпы размножения, чем если бы она размножалась многократно, как все нормальные пальмы.
Кроме того, непонятно, как эта пальма там вообще оказалась. Ведь её ближайшие родственники растут на Аравийском полуострове, в Китае и Таиланде. То есть, на расстоянии 6000 км от этой пальмы. Таким образом, эта пальма не только опровергает дарвинизм, но еще и создаёт очередную биогеографическую загадку. Добавляющуюся в длинный ряд таких же биогеографических загадок, о которых свидетели Дарвина предпочитают вообще не упоминать, когда рассказывают неподготовленным людям о так называемых «биогеографических доказательствах эволюции». То есть, дарвинисты-проповедники «забывают» упомянуть, что на каждое «биогеографическое доказательство эволюции» существуют свои «биогеографические загадки», которые необходимо специально и дополнительно «объяснять», чтобы и они тоже смогли вписаться в рамки «единственно верного учения». Причем некоторые «загадки» всё равно не вписываются, то есть, удовлетворительных объяснений вообще нет. Но об этом позже.
Еще один теоретический пример, где мы могли бы наблюдать молниеносные эволюционные изменения, можно привести в отношении бабочек или каких-нибудь жуков. Как известно, эти насекомые развиваются, используя полное превращение. Поэтому всю их жизнь можно разбить на стадии яйца, личинки, куколки и взрослой особи. Давайте представим себе, что будет, если какое-то насекомое вдруг решит размножаться на стадии гусеницы, а не взрослой особи.
Допустим, смертность насекомых на стадии яиц – 95%, на стадии гусеницы – еще 95%, на стадии куколки - 50%, и наконец, на стадии взрослого насекомого – 20%. И допустим, у какой-то отдельной особи произошла мутация, в результате которой всё потомство этого насекомого дружно «решило» начать размножаться уже на стадии гусеницы (вместо того, чтобы окукливаться). В этом случае, гусеницы, которые выйдут из яиц этой мутантной особи и доживут до размножения (именно на стадии гусеницы) - внесут совокупный вклад в следующее поколение (в виде яиц) в 2.5 раза выше, чем нормальная (не мутантная) особь, потомство которой (зачем-то) «терпит» с размножением до стадии взрослого насекомого (через куколку), резко теряя при этом в численности за счет гибели на дополнительных стадиях. Это обстоятельство особенно интересно, если учесть, что некоторые насекомые на взрослой стадии вообще не питаются. Поэтому и стадия куколки (которая тоже не питается) и стадия взрослого насекомого – обе эти стадии живут только за счет тех резервов, которые накопила гусеница. И эти резервы гусеница могла бы, в принципе, сразу же пустить на производство яиц, не откладывая это дело в долгий ящик. А если мы еще вспомним, что существуют бабочки-долгожительницы (например, лимонницы), которые беззаботно порхают (не торопясь с размножением) в течение целого летнего сезона, а размножаются только на следующий год (!), то смертность таких бабочек вряд ли будет составлять озвученные нами 20%. Скорее всего, в разы больше. И если их смертность достигнет, например, 80%, тогда гусеницы, которые «решились» бы сразу отложить яйца, не дожидаясь стадии своих порхающих бабочек, отложили бы (в совокупности) в 10 раз (!) больше яиц, чем все взрослые бабочки, сумевшие дожить до размножения.
Первое, что здесь напрашивается - что взрослая стадия нужна этим насекомым для расселения, поскольку только эта стадия умеет летать (а гусеница не умеет). Но на самом деле, биологическая целесообразность полёта тоже находится под вопросом. Потому что на полёт тоже нужно расходовать ресурсы. И если некий биологический вид обитает в стабильных местообитаниях, которые обширны и непрерывны, то летать ему там особенно некуда. Ибо куда бы он там ни полетел, он везде встретится с другими такими же особями своего вида. Поэтому вопрос, что же всё-таки лучше – куда-нибудь полететь, или же вообще никуда не летать, а на сэкономленные ресурсы произвести большее число яиц – в таких условиях этот вопрос весьма актуален. И целый ряд нелетающих видов бабочек поступает именно вторым образом. Например, большинство видов мешочниц. У мешочниц самки часто не только бескрылые, но еще и нередко безногие, и размножаются они, не выходя из кокона. Или, например, бескрылые самки зимней пяденицы, или пяденицы-обдирало – тоже никуда не летают. Или полностью крылатые, но всё равно никуда не летающие самки непарного шелкопряда (европейского подвида), и так далее.
Например, самки тропической бабочки Hypolimnas misippus, несмотря на то, что умеют летать (обладая в совершенстве развитыми красивыми крыльями), тем не менее, предпочитают выбирать местечки для откладки яиц методом обычной прогулки (ногами), а не с помощью полета. Эту особенность их поведения биологи склонны объяснять именно экономичностью данного способа передвижения (Atluri et al., 2013). Возникает вопрос – что же тогда мешает совершать такие же прогулки (для откладки яиц) уже на стадии гусеницы? [99]
Итак, где же в природе такие виды бабочек, которые в ходе аналогичной эволюции стали, «не мудрствуя лукаво», просто откладывать яйца уже на стадии гусеницы? Неужели нашей неисповедимой эволюции выполнить такую задачу оказалось нереально сложно? То есть, оказалось невозможным просто удалить (за ненадобностью) стадию куколки хотя бы у некоторых видов (из 150 тысяч известных видов бабочек)… хотя с другой стороны, эта же эволюция как-то сумела (в воображении верующих дарвинистов) создать и яйца, и гусениц, и куколок, и взрослых бабочек.
Ну и пожалуй, следует завершить наши изыскания таких примеров в живой природе, где мы могли бы (в принципе) наблюдать молниеносное распространение соответствующих признаков в популяциях - примером, где необходимость «неотложной эволюционной помощи» просто вопиёт.
Этот пример – печально знаменитая половая система пятнистых гиен. Предоставляю слово автору научно-популярной книги (Джадсон, 2012):
…Внешне гениталии самцов и самок вашего вида настолько похожи, что пятнистых гиен долгое время считали гермафродитами. Однако то, что у самок принимали за фаллос, на самом деле - чудовищно крупный клитор, полностью способный к эрекции. Половые губы у них сросшиеся, из них сформирована так называемая псевдомошонка. Таким образом, мочеиспускание, совокупление и рождение детенышей происходит через клитор.
Каким образом? Ну, если ты действительно хочешь знать. В пубертатном возрасте створ клитора становится эластичным и способным открываться примерно на два сантиметра в диаметре. При половом сношении самка втягивает клитор, складывая его гармошкой, и в открывшееся отверстие проникает пенис партнера.
Однако особенно необычно проходит у пятнистых гиен рождение детенышей. Неискушенным родовой канал гиены покажется крайне странным по форме. Вместо того чтобы как у других млекопитающих, вести непосредственно наружу он резко изгибается. Более того, он достигает 60 сантиметров в длину - вдвое больше, чем у других млекопитающих сходного размера. При этом пуповина у них очень короткая - едва достигает 18 сантиметров. Роды должны быть быстрыми, иначе после отделения плаценты юная гиена умрет от удушья. Однако голова детеныша слишком велика, чтобы пройти через клитор, поэтому у впервые рожающей гиены клитор разрывается, чтобы дать потомству возможность выбраться наружу. Это не просто больно, но зачастую смертельно опасно. По подсчетам ученых, примерно 10% самок пятнистой гиены умирают при первых родах, при этом около половины детенышей у впервые рожающих самок появляются на свет мертвыми. (Парадоксально, что, поскольку клитор не восстанавливается после травмы, дальнейшие роды проходят без всякого риска для матери.).
Итак, что мы имеем? Мы имеем просто поразительный набор признаков, которые в совокупности очень вредны для пятнистой гиены. Во-первых, такие особенности их половой системы приводят к 10%-ной смертности взрослых самок, а во-вторых, от этого умирает еще и около половины первого потомства. А ведь мы с Вами только что (!) рассуждали о том, насколько первое потомство может быть важнее любого другого.
Но пожалуй, самым поразительным здесь является то обстоятельство, что не только у всех остальных млекопитающих, но даже у самых близких к пятнистой гиене биологических видов (бурая гиена, полосатая гиена, земляной волк) нет ничего подобного тому «чуду-юду», которое мы наблюдаем у пятнистых гиен.
Понятно, что если бы в популяции пятнистых гиен в какой-то момент появилась отдельная мутантная особь с нормальной половой системой, то за счет 10%-ной разницы в выживаемости, потомки такой особи вытеснили бы немутантных гиен всего примерно за 100-150 поколений. А поскольку к этому добавилась бы еще и повышенная выживаемость первого потомства, то скорость подобной замены могла бы стать еще быстрее. То есть, практически мгновенной по эволюционным меркам.
Однако пятнистая гиена... продолжает оставаться пятнистой гиеной.
Более того, совершенно непонятно, каким образом признак, столь опасный для жизни, вообще завоевал популяции пятнистых гиен в ходе воображаемой «эволюции более приспособленных». Скорее уж, здесь следует предположить нечто совершенно другое – что половую систему гиен создавала такая сила, которая, наоборот, задалась специальной целью максимально осложнить гиенам их размножение. Или решила создать животных с таким специфическим стилем дизайна, где чуть ли не в каждой видовой особенности сквозит жестокость и кровожадность. Начиная от внешности, и заканчивая половым и охотничьим поведением. И даже спецификой взаимоотношений между детьми в потомстве.
К сожалению, верующие дарвинисты крайне далеки от понимания, что живые существа могут выглядеть так или иначе… просто потому, что у них такой стиль дизайна (т.е. просто ради разнообразия). Поэтому они пытаются как-то объяснить столь удивительный набор черт в половой системе гиен в рамках своего любимого учения. Но поскольку в рамках их любимого учения, сложные признаки можно объяснять, только придумывая им какую-нибудь биологическую полезность, то с пятнистыми гиенами дело пока идёт туго. Точнее, вообще никак не идет. Продолжаю цитату (Джадсон, 2012):
Итак, мы имеем странный набор фактов. Гиенам, как мы предполагаем, досталось два эволюционных преимущества. Первое - наличие органа, имитирующего фаллос, имея который самки могут участвовать в приветственных церемониях: при встрече двух пятнистых гиен они утыкают носы в хвосты друг друга и изучают эрегированные члены друг друга. Участие самок в этих церемониях, вполне вероятно, помогает им удерживать свое господство над самцами. Тем не менее, хотя эта идея наверняка понравится фрейдистам, едва ли она способна объяснить наличие такого опасного органа.
Следующее предположение еще более умозрительно. Механизм секса у пятнистых гиен столь сложен, что самка всегда сможет противостоять непрошеному ухажеру: соитие возможно лишь при доброй воле обеих сторон, изнасилование невозможно в принципе. Однако ни у одного вида гиен ученые до сих пор не зафиксировали случаев сексуального насилия. Более того, поскольку самки пятнистой гиены значительно крупнее самцов, к тому же обладают мощными челюстями, ухажеры у этого вида отличаются небывалой учтивостью: они приближаются к избраннице, кланяясь буквально до земли. Так что фаллос вряд ли нужен самкам для самозащиты.
Думаю, вы согласитесь: ни одно из этих объяснений нельзя считать удовлетворительным. А может быть, фаллос - лишь побочный продукт естественного отбора, имевшего совершенно иную цель? На первый взгляд, эта идея кажется более правдоподобной. Есть и кандидат на роль «иной цели»: агрессия. Мы знаем, что гиены агрессивны, и можем предположить, что злобные самки оказываются более успешными, нежели тихони. Более того, зародыш гиены, еще находясь в матке, получает большие дозы тестостерона и других андрогенов - мужских половых гормонов. Внутриматочное воздействие этих гормонов пробуждает агрессивность: так, самки мыши, зажатые в материнской матке между своими братьями и поэтому получавшие более заметные дозы андрогенов, во взрослой жизни оказывались более агрессивными, нежели их сестры, которым досталось место в окружении девочек. При этом внутриматочное воздействие больших доз андрогенов может вызывать существенные отклонения в строении гениталий. К примеру, у людей, если девочка в материнской утробе получает избыточные дозы этих гормона, она рождается с сильно увеличенным клитором и частично сросшимся влагалищем. Таким образом, возникает вопрос: может ли естественный отбор столь сильно благоприятствовать развитию агрессии у самок пятнистой гиены, чтобы совокупление и роды через клитор оказались не слишком высокой ценой?
В принципе, может. Пятнистые гиены рождаются такими, какими им предстоит провести большую часть жизни, - с оскаленными зубами. У большинства гиен щенки рождаются по двое и тот, который появился на свет первым, тут же набрасывается на собрата. В результате один из щенков зачастую гибнет. Убив брата или сестру, можно получить монополию на материнское молоко; поскольку гиены кормят щенков грудью более года, убийство второго претендента повышает шансы дожить до зрелости. Действительно, одна из теорий заключается в том, что высокий уровень андрогенов в матке обеспечивает преимущества именно потому, что он провоцирует насилие среди щенков при рождении. Правда, эта версия не объясняет, почему убийства часто случаются в однополых парах, чем между братьями и сестрами и почему самки убивают друг друга чаще, чем самцы. Если бы дело было просто в братоубийстве, пол соперника не имел бы значения.
Более убедительное объяснение гласит, что агрессия важна для вида, поскольку именно она регулирует отношения доминирования, а доминантный лидер пользуется немалыми преимуществами. Высокопоставленные самки в сравнении с теми, кто стоит ниже в социальной иерархии, раньше беременеют, чаще рожают, а их потомки имеют больше шансов дожить до зрелости. Это - серьезное преимущество, которое может стоить неудобств, связанных с фаллообразным клитором.
Тем не менее загадка не решается столь легко. Исследования пятнистых гиен показывают, что блокирование поступления андрогенов в матку не приводит к возвращению к «типичным» гениталиям самок. Получается, что развитие фаллического клитора по большей части не зависит от этих гормонов, что подрывает нашу теорию, гласящую, что женский фаллос – побочный продукт естественного отбора, направленного на повышение агрессивности. Так что, пока мы не узнаем больше о том, за счет чего развивается этот орган, боюсь, причина, по которой ты наделена столь необычным и доставляющим столько неприятностей отростком, так и останется загадкой.
Итак, было высказано предположение, что самки гиен имеют столь поразительную половую систему потому, что у них развитие идет по мужскому типу - от переизбытка мужских гормонов женская половая система начинает развиваться в соответствующую сторону. Но оказалось, что от концентрации андрогенов формирование удивительных особенностей женской половой системы гиен не зависит.
В связи с этим можно предположить, что псевдо-мужские половые органы самок гиен – это, на самом деле, не побочный результат деформирования женских половых органов к мужскому типу, а специальные морфологические черты, которые имитируют мужские половые органы (чисто внешне). И верующие дарвинисты пока не знают, зачем это понадобилось пятнистым гиенам. Однако подсказок от креационистов они принимать не хотят.
Хочу добавить, что автор цитаты говорит о повышенном уровне агрессии гиен, как о безусловно полезном признаке. Хотя такое утверждение, как минимум, спорно. Точно так же (чисто умозрительно) можно заявить, что повышенная агрессивность гиен, наоборот, вредит и этому виду в целом, и отдельным особям. И после этого пригласить рассказать что-нибудь на эту тему Ричарда Докинза. Ведь Ричард Докинз в своё время любил порассуждать на тему «благородства» животных, доказывая, что когда животные одного вида сдерживают агрессию друг против друга, то это им, на самом деле, биологически выгодно.[100] Таким образом, верующие дарвинисты ведут себя, как профессиональные гадатели на кофейной гуще, которые видят в кофейной гуще каждый раз именно то, что им удобно видеть в рамках «единственно верного учения». И когда эти гадатели начинают говорить прямо противоположные вещи, их «очная ставка» была бы весьма поучительной.
Пусть бы они вместе с Оливией Джадсон (автором приведенного отрывка про гиен) обсудили бы этот вопрос. Со стороны это будет здорово смотреться. Один убежденный дарвинист будет уверять, что повышенная агрессия – полезна для этого вида, а другой убежденный дарвинист будет говорить, что полезна пониженная агрессия. Точь в точь профессиональные гадатели.
Ну а на самом деле, мы знаем много разных видов зверей, самки которых вполне достаточно агрессивны (достаточно для успешного выживания этого вида). Начиная с домашних кошек, и кончая росомахами, тигрицами или медведицами. Мы не можем назвать этих самок «мирными одуванчиками». Тем не менее, никто из них не рождает детенышей через клитор. И их дети не убивают друг друга вскоре после рождения. Даже такие социальные звери (со сложной системой иерархии), как волки или гиеновидные собаки - тоже не имеют ничего подобного тому, что демонстрируют нам пятнистые гиены.
Однако чего это нас всё время тянет опровергать дарвинизм? Ведь мы в этой главе просто считаем рейтинги скорости (воображаемой) эволюции в зависимости от силы давления естественного отбора. А мы явно увлеклись описанием особо экстравагантных «чудес природы». Начиная с бамбука, погибающего после первого же размножения семенами, и кончая поразительными самками пятнистых гиен, половая система которых зачем-то имитирует половую систему самцов. Причем внешнее сходство, действительно, впечатляет. Чего никак нельзя сказать об эффективности размножения обладательниц этого «чуда природы».
Увлеклись мы в том смысле, что вряд ли, например, переход каких-нибудь бабочек к размножению на стадии гусеницы – это такое уж легкое эволюционное изменение, которое может произойти благодаря одной единственной мутации. Скорее всего, для такого события требуется внести целый ряд генетических изменений. А раз так, то можно сказать, что подобные изменения могут появляться в природе либо очень редко, либо очень постепенно (правда, тогда не очень понятна польза от такой «постепенности»). И следовательно, мы и не наблюдаем сегодня в природе подобных изменений.
Хотя я старался подбирать такие примеры, которые косвенно свидетельствуют именно о возможной легкости соответствующих эволюционных изменений… в рамках самой же теории эволюции. Действительно, существование в пределах одного и того же рода рыб (Oncorhynchus): 1) «одноразовых» лососей, 2) «многоразовых» лососей и 3) лососей, в целом, «одноразовых», но у которых отдельные особи могут быть «многоразовыми» - существование такого рода рыб как бы намекает на то, что подобные изменения должны осуществляться легко. Если мы, конечно, считаем, что эти различия являются именно результатом эволюции, а не чего-нибудь другого. Или, например, существование многочисленных линий организмов, размножающихся с помощью партеногенеза в рамках одних и тех же таксонов с видами, которые размножаются «классическим» половым способом - начиная от разных насекомых и кончая рыбами, амфибиями и ящерицами – этот факт тоже намекает нам, что такие эволюционные переходы могли бы осуществляться достаточно легко. Наконец, те же пятнистые гиены, в рамках всё той же теории эволюции, шли к своей поразительной половой системе (вот только зачем шли?) тоже, наверное, не слишком тяжелыми эволюционными путями. Потому что ни один из ближайших родственников пятнистых гиен ничего подобного не имеет.
Тем не менее, чисто теоретически, возражение насчет возможной сложности (и следовательно, редкости подобных изменений в природе) в данном случае привести можно. Поэтому давайте на всякий случай зафиксируем скорость распространения некоего (теоретического) признака, повышающего приспособленность организмов вдвое, и дальше сосредоточимся уже на таких признаках, для реализации которых было бы достаточно всего лишь одной мутации - в прямом смысле этого слова.
1. Итак, если какой-то признак повышает приспособленность особи на 100%, то он завоюет популяцию всего примерно за 20 поколений. Запомним это.
2. Если же какой-то признак повышает приспособленность на 10%, то, как я уже говорил выше, этот признак может завоевать популяцию всего примерно за 150 поколений. Даже для полевых мышей, которые размножаются 3-4 раза в год, эти 150 поколений реально воплощаются в 40-50 лет. То есть, эволюцию подобных признаков мы могли бы пронаблюдать прямо на глазах (!) даже у позвоночных животных. А не только у березовой пяденицы.
Причем «эволюция березовой пяденицы» нам уже на практике показала, как это могло бы быть в живой природе – небольшое генетическое изменение, но оказавшееся полезным в рамках дополнительных 10% приспособленности, и поэтому лавинообразно распространяющееся в популяциях всего за 40-50 лет (см. ниже).
И вот мы (точнее, дарвинисты) предполагаем, что вся живая природа у нас эволюционирует прямо сейчас под действием естественного отбора. То есть, все существующие миллионы биологических видов (и миллиарды популяций)...
А где примеры?
3. Если какой-то признак повышает приспособленность особи всего на 1%, то он будет завоевывать популяцию примерно в 10 раз дольше, чем предыдущий (который повышал приспособленность на 10%). То есть, такой признак завоевал бы популяцию (до 95% носителей этого признака) примерно за 1000-2000 поколений (а если повезет, то и за 800).[101] Понятно, что 1000-2000 поколений – это уже достаточно серьезный промежуток времени.
Но во многих случаях даже такие эволюционные изменения вполне можно было бы зафиксировать. Во-первых, для многих организмов – тысяча поколений - это всего примерно 250 - 350 лет. Например, для тех же полевых мышей (см. выше). Понятно, что 250–350 лет, в принципе, доступны для прямого изучения, особенно если произошла морфологическая эволюция каких-нибудь косточек, панцирей, раковин, семян или орехов, которые способны легко сохраниться на протяжении озвученного промежутка времени. Или даже листьев и травянистых стеблей, которые вполне могут «протянуть» это время в состоянии гербария. Во-вторых, совсем не обязательно дожидаться полной победы нового признака в популяции. Например, если новому признаку повезёт, то примерно через 400 поколений уже половина особей этой популяции будет обладать этим признаком. Понятно, что когда сначала этого признака не было вообще, а через некоторое время этот признак имеется уже у половины всех встречающихся особей данного вида, то такое обстоятельство вполне могло бы быть замечено даже обычными людьми, не говоря уже о профессиональных биологах. А ведь 400 поколений для тех же полевых мышей – это всего 100-130 лет. И это (напоминаю) при давлении естественного отбора, равном всего 1%. Теоретически, появление таких признаков, которые приводят к разнице в приспособленности всего 1%, кажется вполне вероятным, или даже весьма вероятным. Тем более, если в нашем распоряжении все миллионы биологических видов Земли, из которых, как минимум, десятки тысяч видов уже очень хорошо известны и изучены.
Я уж не говорю о том, что многие виды способны размножаться не по три, а по десять раз в год. Поэтому даже 1000 поколений для таких видов превращается всего в 100 лет.
В общем, проблема здесь не в скорости естественного отбора, которая, на самом деле, весьма быстра и поэтому совершенно не является проблемой для наблюдения «эволюции в действии». Проблема здесь в том, чтобы дождаться появления именно такой случайной мутации, которая окажется полезной для особей того или иного вида, повысив их приспособленность хотя бы на 1-10%. Кроме того, такой мутации (даже если она случится), должно еще повезти на самом начальном этапе. Когда особей, несущих эту мутацию, еще очень мало, и поэтому все они могут легко «вылететь» из популяции вследствие чисто случайных причин (дрейф генов).
Однако с другой стороны, у нас в распоряжении сотни миллиардов особей самых разных живых существ, относящихся к миллионам биологических видов (которые производят на свет многие триллионы или даже биллионы яиц, спор, личинок и детенышей). Что само по себе подразумевает весьма обширный биологический материал для периодического возникновения только что описанных случайных мутаций. Таким образом, теоретически, мы должны были бы, наверное, пронаблюдать уже много таких «эволюционных вспышек», когда какой-либо новый полезный признак внезапно появлялся в той или иной популяции того или иного биологического вида. И потом следовал быстрый эволюционный «захват» этой популяции данным признаком, в соответствии с теми рейтингами скоростей естественного отбора, которые мы здесь уже успели озвучить.
Но что-то никаких «эволюционных вспышек» мы не наблюдаем. Точнее, иногда всё-таки наблюдаем (в единичных случаях). Но в подавляющем большинстве таких случаев после проведения специальных исследований оказывается… что механизмы произошедших изменений были не дарвиновскими (см. ниже).
4. Наконец, если какой-то признак приводит к повышению приспособленности всего на 0.1%, то здесь для практически полной победы этого признака (до 95% носителей) требуется уже примерно 8000–20000 поколений. То есть, эволюционный «захват» популяции подобным признаком уже вряд ли можно будет пронаблюдать непосредственным образом. Кроме того, подавляющая часть таких признаков должна удаляться из популяций чисто случайным образом в самом начале своего появления, за счет дрейфа генов.
И тем не менее, если эти признаки относятся к таким, которые хорошо сохраняются в виде останков, вполне можно было бы зафиксировать изменение и таких признаков тоже. То есть, эволюционные изменения раковин, костей, панцирей, семян и орехов, коры и древесины (и даже в некоторых случаях мумий) вполне можно было бы зафиксировать даже в случае эволюции таких признаков, которые создают разницу в приспособленности особей всего 0.1%.
Если бы, конечно, современная теория эволюции была правдой.
То есть, что можно было бы сделать? Можно было бы измерить сегодняшнее давление естественного или полового отбора по отношению к какому-нибудь признаку вида. И если будет установлено, что это давление не равно нулю, то можно было бы посмотреть, как изменялся этот признак у данного вида в течение последних тысяч или десятков тысяч лет. И всё, что нам останется, это зафиксировать соответствующие изменения данного признака (в соответствующую сторону) в ископаемых останках этого организма (если, конечно, данный признак вообще сохраняется в ископаемом состоянии). Таким образом, с одной стороны мы будем иметь давление естественного (или полового) отбора, строго установленное сегодня, а с другой стороны, установленные эволюционные изменения этого признака именно в ожидаемом направлении, зафиксированные в ископаемых останках. Понятно, что это будет уже не самое прямое доказательство эволюции под действием естественного отбора (из всех возможных). Но всё же такие установленные факты были бы очень мощным свидетельством в пользу этой теории.
Приведу конкретный пример. В ходе целого ряда исследований было установлено, что у женщин Homo sapiens имеются серьезные половые предпочтения по отношению к такому признаку мужчины, как мужская атлетичность (Weeden & Sabini, 2005).[102]
Таким образом, мужская атлетичность должна являться серьезным фактором полового отбора.
Более того, мужская атлетичность приветствуется не только женщинами Homo sapiens, но еще и является важной чертой социальных взаимодействий между самими мужчинами. То есть, сами мужчины очень уважают мужскую атлетичность. По сути, это один из самых первых признаков, которые один мужчина оценивает в другом мужчине. И если атлетичность другого мужчины оставляет желать лучшего, то мы успокаиваемся (и даже можем преисполниться чувством собственного превосходства). А вот если атлетичность другого мужчины явно «выпирает» из всех углов (если фигура явно мощная), то мы начинаем чувствовать себя не совсем комфортно. Причем такое уважение (мужчин) к физической силе и атлетичности наблюдается уже в подростковом возрасте.
Таким образом, атлетичность даёт большое преимущество не только в процессе «завоевания» самки, но и во взаимодействии с другими мужчинами.
Наконец, в отличие, например, от «хвоста» павлина, атлетичность мужчины нельзя назвать вредным признаком. Если хвост павлина откровенно затрудняет выживание павлину и является явно вредным признаком, то атлетичность мужчины имеет не только свои минусы (больше ресурсов необходимо для её поддержания), но и весьма неплохие плюсы - атлетичность позволяет легче справляться с разнообразными физическими нагрузками, в том числе, сопровождающимися риском для жизни.
Итак, по сути, мы имеем признак, который должен эволюционировать в популяциях людей под мощнейшим давлением полового отбора, производимым сразу в двух его формах - и за счет предпочтения самок, и за счет преимущества в конкуренции с другими самцами.
Теперь нам осталось только раскопать несколько десятков скелетов мужчин, начиная от современности и до «глубины» в несколько тысяч (или десятков тысяч) лет. И продемонстрировать, как этот признак (мужская атлетичность) постепенно эволюционировал в соответствующем направлении под мощным давлением полового отбора.
Такая работа уже была проделана. И вот что оказалось. Даём слово выдающемуся специалисту по эволюции человека (автору книг «Эволюция человека» в двух томах) доктору биологических наук, Александру Маркову: [103]
...На самом деле, кстати, наоборот - за последние тысячелетия, по антропологическим данным, люди, по крайней мере многие человеческие популяции, стали менее атлетичными.
Итак, мы видим, что «эволюционные изменения» действительно произошли. Но к сожалению, в совершенно противоположную сторону. То есть, рассуждения в рамках теории полового отбора говорили нам, что мужская атлетичность должна развиваться в ходе эволюции. А она, на самом деле, оказывается, деградировала.
То есть, вместо «доказательства эволюции по механизму полового отбора», мы получили очередной и совершенно замечательный пример, насколько на самом деле «справедлива» теория полового отбора, и как «замечательно» сбываются её прогнозы.
Интересно, что имеется забавное опровержение ламаркизма (в смысле наследования приобретенных признаков) тоже на примере людей. Как известно, девственная плева «вынужденно не используется» в течение жизни подавляющим большинством женщин. Тем не менее, девственная плева продолжает (в норме) сохраняться в популяциях людей. Этот факт наглядно опровергает знаменитое ламарковское «наследование приобретенных признаков».
Точно так же и здесь – мужской атлетизм крайне важен для полового отбора (причем сразу для обеих его форм). Но он не только не развивается в ходе эволюции, а наоборот, постепенно деградирует по факту.
Тем не менее, никто почему-то не говорит, что этот четкий факт опровергает эволюцию под действием полового отбора в природе (в отличие от девственной плевы, где все согласны, что она ламаркизм опровергает). Вместо признания опровержения теории полового отбора (по факту) верующие дарвинисты в этом случае начинают приводить самые разные оправдания, выполненные в стиле:
– Почему так получилось, что опять ничего не получилось.
То есть, начинаются вольные размышления о том, что же так помешало бедным мужчинам развить себе атлетичность под действием полового отбора. Причем помешало настолько, что у них вообще ничего из этого не вышло, несмотря на совершенно четкие женские и мужские предпочтения именно к этому признаку.
Как правило, в качестве подобной «эволюционной помехи» озвучивается ограниченность пищевых ресурсов. И это у человека! Интересно, если даже человек, который сам производит себе пищевую продукцию (причем как растительную, так и мясную), оказывается, настолько ограничен в пищевых ресурсах, что это начисто заблокировало его предполагаемый половой отбор, то что уж тогда говорить обо всех остальных биологических видах с ярко выраженными «архитектурными излишествами», которые никакой пищи себе не выращивают, а выживают в дикой природе, где по уверениям самих же верующих дарвинистов имеется страшный дефицит ресурсов. Как же эти биологические виды умудрились выделить дополнительные ресурсы на реализацию столь роскошных «половых излишеств», некоторые из которых могут превышать длину всего остального тела этого животного? [104]
Понятно, что придумать какое-то «объяснение» задним числом – в этом у дарвинизма просто нет равных. Хоть как-то конкурировать с дарвинизмом здесь может, наверное, только астрология, гадание на кофейной гуще, и еще фрейдизм. Эта особенность дарвинизма всем хорошо известна, являясь уже «притчей во языцех». Вот, например, как иронично прокомментировал эту ситуацию один из участников обсуждения этой темы:
Еще раз повторяем цитату Александра Маркова:
...На самом деле, кстати, наоборот - за последние тысячелетия, по антропологическим данным, люди, по крайней мере многие человеческие популяции, стали менее атлетичными. Потому что женские предпочтения по этому признаку - лишь один из многих факторов отбора.
Встречный комментарий читателя: [105]
…Что, в принципе, не помешает в будущем предположить, что … женщины полюбили и стали отбирать … астеничных мужчин.
Если же всякие теоретические спекуляции, выполненные задним числом, вообще не рассматривать, а брать только голые факты, то по факту мы имеем следующее. Вот что пишет антрополог С.В. Дробышевский о скелете мужчины с широко известной палеолитической стоянки древнего человека Сунгирь (около 25 тысяч лет назад): [106]
…Последний индивид имел чрезвычайно широкие плечи при довольно высоком росте и сравнительно узком тазе. Своей массивностью он выделяется даже среди других верхнепалеолитических сапиенсов.
Так вот, «широкие плечи при довольно высоком росте и сравнительно узком тазе» - это как раз и есть те самые классические признаки мужской привлекательности, от которых так млеют наши женщины.[107] Таким образом, в самом начале нашей истории мы уже имели практически идеал «женского полового отбора». А вот к настоящему времени этот идеал заметно «поистрепался», несмотря на неусыпный «половой отбор» со стороны наших женщин на протяжении всех прошедших 25 тысячелетий.
Кстати, можно немного поиронизировать над верующими дарвинистами еще и по такому антропологическому поводу. Известно, что сравнительно недавно антрополог Лавджой буквально потряс всех новой «теорией» происхождения человека из обезьяны. Если раньше Фридрих Энгельс провозгласил довольно красивый лозунг про то, как:
- Труд сделал из обезьяны человека.
То антрополог Лавджой провозгласил совершенно новую движущую силу антропогенеза:
- Проституция сделала из обезьяны человека.
По мнению Лавджоя, древние человекообразные обезьяны встали на задние лапы, возможно, потому, что самцам пришлось носить в передних лапах еду своим самкам. Поскольку эти самки стали придерживаться принципа «секс в обмен на пищу» (Lovejoy, 2009).
И вот в результате такого полового отбора,[108] через много-много лет и развился человек из обезьяны.
Что и говорить, верующие дарвинисты, как говорится, «отжигают». Причем эти вольные рассуждения Лавджоя (с некоторыми признаками сексуальной озабоченности) редакция журнала Science назвала… «научным прорывом 2009 года»!
Так и хочется ответить на это словами одного мультипликационного персонажа:
- Да уж, это прорыв… (С) [109]
Никак не меньше отожгли и другие авторы, которые предположили, что:
- Бокс сделал из обезьяны человека.
Они даже проделали серию экспериментов, в которых выяснили, что наиболее хороший удар кулаком получается в том случае, если рука имеет такое же строение, как человеческая кисть. Вот авторы и предположили, что лапы обезьян эволюционировали в человеческие кисти (хотя бы частично) в результате боксерских матчей, проводимых между самцами этих обезьян. То есть, в результате соответствующего полового отбора (Morgan & Carrier, 2013).
К сожалению, авторы этой гипотезы не учли, что само сжатие кисти в кулак и правильный удар таким кулаком – это определенное искусство, которому необходимо сначала специально обучиться (прежде чем бить). Иначе рискуешь остаться с выбитыми пальцами.
В общем, создаётся ощущение, что антропологов кто-то специально отбирает на предмет богатства воображения и буйства фантазии. Может быть, это как раз и есть тот самый долгожданный пример естественного отбора?
На самом деле, вопрос, что же побудило древних обезьян встать на задние лапы, похоже, уже достаточно ясен. Скорее всего, ведующую роль в ранних этапах эволюции человека из обезьяны сыграла аллергия. Дело в том, что в то время стали происходить климатические изменения, которые привели к расширению саванн и сокращению лесов. Поэтому в воздухе появилось, во-первых, существенно больше пыли, а во-вторых, появилась аллергенная пыльца тех видов травянистых растений саванны, к которым древние человекообразные обезьяны были непривычны. В связи с этим у древних обезьян стали чесаться носы, и во многих случаях развивалась аллергия разной степени тяжести. Отчего древние обезьяны впадали в стресс и начинали хуже размножаться. В этих условиях преимущество в выживании получали те особи, которые могли чаще вставать на задние лапы, освобождая руки для того, чтобы почесать у себя в носу. Именно те обезьяны, которые могли чаще и дольше ковыряться в носу пальцами, стали выигрывать борьбу за существование у своих менее одаренных сородичей. Под давлением этого нового фактора естественного отбора, начался соответствующий эволюционный процесс, который, в конце концов, и привел к тому, что древние обезьяны совершенно освободили руки, став полностью двуногими.
Поскольку подобную гипотезу становления двуногости у древних приматов еще никто не озвучивал, то я в ближайшее время собираюсь опубликовать эту гипотезу в том же самом журнале, где антрополог Лавджой опубликовал свою. Может быть, моя гипотеза тоже будет названа новым «научным прорывом»? Тем более что моя гипотеза, в отличие от гипотезы Лавджоя, прекрасно объясняет, почему на задние лапы встали сразу оба пола обезьян, а не только самцы (как следовало бы ожидать из гипотезы Лавджоя).
В ожидании же этой эпохальной научной публикации давайте пока возвратимся именно к версии Лавджоя - к эволюции человека из обезьяны в результате «секса за еду». Многие глубоко верующие дарвинисты с энтузиазмом восприняли эту гипотезу. Но дело в том, что эволюция по механизму «секс в обмен за пищу» относится к области именно полового отбора.[110] Таким образом, получается, что именно половой отбор сумел создать из какой-то «древней гориллы» – человека.
И в то же самое время, когда спрашиваешь дарвиниста – почему мужчины не развили себе атлетичные фигуры, несмотря на половой отбор, дарвинисты отвечают, что мужчины просто «не шмогли». Потому что не хватало ресурсов для такой эволюции. Тем не менее, той же горилле вполне хватает ресурсов (в дикой природе!), чтобы набрать массу тела, равную примерно 250 кг, имея при этом фигуру исключительной атлетичности.
Давайте посмотрим для наглядности на современную гориллу (Рис. 34):

Рисунок 34. Самец гориллы во всей красе.
Наверное, многие мужчины мечтали бы иметь такие мышцы спины, рук и плеч, как современная горилла. Тем не менее, они их не имеют. Несмотря на то, что питаются (в том числе) мясом, причем мясо нередко достигает значительной доли в рационе. А вот горилла нарастила себе такие мышцы почти исключительно на вегетарианской диете.
Таким образом, получается, что верующие дарвинисты в этом конкретном случае просто не верят в идею «эволюции без границ». Они не верят, что человек под действием полового отбора смог бы превратиться в гориллу (и выдвигают разные воображаемые ограничивающие факторы). И в то же самое время, эти же загадочные дарвинисты почему-то верят, что какая-то (древняя) горилла смогла превратиться в человека… под действием всё того же полового отбора. Что и говорить, завидовать логическому мышлению верующих дарвинистов вряд ли стоит.
На самом деле, придумывая какие-то очередные объяснения, почему мужчины Homo sapiens под действием мощнейшего (двойного) полового отбора, тем не менее, «сэволюционировали» не в сторону повышения атлетичности, а наоборот, в сторону её деградации… верующие дарвинисты фактически признают, что у биологического вида Homo sapiens имеются определенные пределы и по массе тела, и по атлетичности, которые характерны для этого вида, и которые этот вид преодолеть просто не может .
А вот, например, горилла преодолевает эти пределы с легкостью. Зато для неё, наоборот, невозможно стать такой же «стройной», как Homo sapiens. То есть, получается, что у гориллы имеются уже свои границы этих признаков, характерные уже для этого биологического вида, которые не совпадают с границами этих же признаков, характерных для человека. И похоже что эти (видовые) признаки просто не могут эволюционировать, то есть, не могут преодолевать рамки своих возможных значений (для этого вида) несмотря ни на какие (даже самые мощные) «половые предпочтения».
Во всяком случае, мы теперь имеем хороший пример, как под действием строго установленных половых предпочтений соответствующая эволюция не состоялась. И одновременно не имеем (пока) ни одного строго установленного примера, где бы такая эволюция вообще состоялась. В связи с этим, меня даже терзают сомнения – а стоило ли вообще терять время на написание книги, опровергающей существование некоего воображаемого явления, если это явление вообще не зафиксировано в реальной природе?
Однако мы опять незаметно ушли от обсуждения возможных скоростей эволюции (под действием естественного или полового отбора), отвлекшись на буйно-фантастические гипотезы антропогенеза.
Тем не менее, мы всё-таки успели обсудить то, что хотели. Подводим итоги:
1) Признак даёт преимущество в приспособленности +100% - ожидаемая скорость распространения признака 20 поколений.
2) Признак даёт преимущество в приспособленности +10% - ожидаемая скорость распространения признака примерно 150 поколений.
3) Признак даёт преимущество в приспособленности +1% - ожидаемая скорость распространения признака примерно 1000-2000 поколений.
4) Признак даёт преимущество в приспособленности +0.1% - ожидаемая скорость распространения признака примерно 8000-20000 поколений.
И пришли к общему выводу, что учитывая огромный биологический материал, который имеется в распоряжении биологов (вся живая природа Земли с миллионами видов и миллиардами популяций), биологи вполне могли бы уже (за 150 лет существования дарвинизма) разыскать и накопить целый массив убедительных фактов наблюдаемой эволюции в природе под действием именно естественного или полового отборов… если бы такая эволюция действительно имела место в природе. Но вместо этого массива, мы имеем 12.5 примеров «эволюции», из которых 90% либо установлено недостаточно надежно, либо вообще раскритиковано. Такая картина – уже сама по себе весьма красноречива и является хорошим показателем, насколько, на самом деле, «справедливо» данное учение.
Тем не менее (абстрагируясь от этой печальной ситуации), озвученные рейтинги скоростей эволюции теперь позволят нам примерно ориентироваться в тех примерах «эволюции», которые мы будем разбирать ниже. Например, если в каком-либо «примере эволюции» скорость произошедшей замены признака составила, допустим, 50 поколений, а давление естественного отбора при этом было установлено в районе 10-20%, то такая скорость эволюции примерно совпадает с теоретически ожидаемой. Следовательно, предположение именно о естественном отборе, как причине зафиксированных изменений, в данном случае будет вполне достаточным и убедительным.
А вот если, допустим, измеренное давление естественного отбора оказалось низким, и не превышало, например, 1%, но тем не менее, установленная скорость эволюционных изменений оказалась очень высокой, например, всего 10–20 поколений (вместо необходимых 1000-2000), то отсюда следует, что за эти изменения было ответственно что-то другое, но никак не естественный отбор, который просто не успел бы распространить данный признак за такое короткое время при столь небольшом оказываемом давлении. Давайте запомним это обстоятельство.
Дополнительно помочь здесь может еще и сама картина распространения признака. Например, если сначала признак регистрировался лишь у единичных особей, но постепенно распространился на всю популяцию, то такая картина соответствует именно механизму естественного отбора.
Если же признак взрывообразно появляется сразу у целой массы особей, то здесь можно сказать совершенно определенно, что в этом были повинны не случайные мутации (с последующим отбором), а что-то совершенно другое. И это обстоятельство тоже давайте запомним.
И наконец, что будет, если признак – очень слабо полезный? Например, если признак повышает выживаемость на 0.01%, или на 0.001%, или даже на 0.0001%? В этом случае, признак является почти нейтральным. Теоретически, естественный отбор такого признака всё равно должен идти, но этот отбор растянется уже на миллионы поколений (т.е. практически на неопределенное время). Такие медленные изменения действительно уже весьма трудно пронаблюдать и непосредственно зафиксировать.
Причем здесь возникает еще одна теоретическая трудность. Возникает вопрос, а способна ли эволюция вообще идти по таким признакам, которые повышают приспособленность на столь малую величину? Чисто теоретически (если ничего больше не учитывать), то способна. Но имеем ли мы право в данном случае ничего больше не учитывать?
Во-первых, что такое «миллионы поколений»? За такое время в большинстве местообитаний Земли условия изменятся настолько радикальным образом, что столь вялотекущий отбор вполне может стать уже не актуальным. Причем несколько раз подряд. Например, еще 10.000 лет назад пустыня Сахара была цветущей страной. А 20.000 лет назад в Европе был ледниковый период. Громадный ледник покрывал всю северную её часть. Наверное, только в экваториальной зоне Земли можно допустить существование столь стабильных условий, что даже отбор некоего признака длиной в миллионы поколений успеет успешно завершиться до того, как условия среды поменяются. В других же регионах Земли столь медленная фиксация признаков вряд ли успеет дойти до своего успешного завершения прежде, чем в конкретной экосистеме что-нибудь серьезно поменяется.
Во-вторых, вероятность закрепления столь слабо полезных мутаций в популяциях небольшого размера очень низка (вследствие дрейфа генов в самом начале этого процесса). Например, в популяции со средним размером 1000 особей вероятность успешного закрепления мутации, повышающей общую приспособленность организма на 0.001%, будет всего примерно 0.05%. То есть, только одна (!) из примерно 2000 таких слабо-полезных мутаций вообще сможет закрепиться в этой популяции.
Но в это же время происходят ведь и другие мутации. Причем принято считать, что подавляющее большинство этих мутаций – вредные или слабо-вредные. И если слабо-вредных мутаций в тысячи раз больше, чем полезных мутаций, то получается, что на одну слабо-полезную мутацию, которой удалось закрепиться и победить в популяции, придётся в десятки, сотни, или даже в тысячи раз больше слабо-вредных мутаций, которым тоже удастся закрепиться в этих же популяциях по чисто случайным причинам. А это значит, что любая популяция со временем будет постепенно вырождаться. То есть, общая приспособленность любой популяции должна постепенно падать, вследствие того, что естественный отбор не будет успевать удалять из популяций слабо-вредные (или слабо-слабо-вредные) мутации из того общего потока мутаций, которые будут происходить в этой популяции. В результате, накопление вредных мутаций будет идти быстрее, чем их удаление из популяции. Ситуация еще больше усугубляется, если вспомнить, что значительная часть этих слабо-вредных мутаций будут рецессивными. Следовательно, такие слабо-вредные мутации могут «входить» в популяцию вообще как нейтральные. Более того, в этом случае смогут «войти» и достаточно распространиться даже и не очень то слабо-вредные мутации. А может быть, и просто вредные (без приставки «слабо»).
В общем, озвученная проблема - «слабо-полезные мутации vs слабо-вредные» теоретически весьма актуальна. Возникает вопрос, а возможна ли вообще столь медленная эволюция по слабо-полезным признакам в рамках механизма естественного отбора? Или же эволюция под действием естественного отбора может идти только по таким признакам, которые повышают приспособленность особей достаточно ощутимым образом? Но тогда и эволюционные изменения по такому сценарию должны протекать быстро. Настолько быстро, что многие такие случаи мы вполне могли бы замечать и фиксировать (если бы теория естественного отбора была правдой).
Интересно, что компьютерное моделирование всех этих процессов в популяциях, учитывающих как естественный отбор полезных мутаций, так и мощный поток слабо-вредных мутаций, показывает, что естественный отбор, скорее всего, не способен даже удержать популяцию на приемлемом уровне приспособленности (Sanford et al., 2007a,b). Приспособленность популяции будет постепенно снижаться из-за постепенного накопления слабо-вредных мутаций, с которыми естественный отбор не будет успевать справляться. Таким образом, если предложенная компьютерная модель верна во всех исходно заданных параметрах, то о теории естественного отбора можно смело забыть. И начинать говорить о постепенном вырождении мира живой природы.[111]
Но компьютерная модель – это всё-таки теоретическая вещь. Может быть, в ней тоже что-то не учтено, или учтено как-то не так. Поэтому опираться на компьютерные модели (пусть даже самые продвинутые) в опровержении дарвинизма мы не будем. А будем опираться только на строго установленные эмпирические факты.
На этом мы заканчиваем с предварительными пояснениями. И переходим к разбору конкретных установленных примеров «наблюдаемой эволюции».
Дата: 2019-02-02, просмотров: 527.