Предыдущий большой раздел текста я посвятил тому, чтобы объяснить (и потом еще много раз повториться), что никакие факты молекулярной филогенетики не могут свидетельствовать в пользу того или иного механизма, которые привели к генетическим изменениям. Была ли это интеллектуальная модификация геномов, или же это был результат естественного отбора случайных мутаций – филогенетика на этот счет уверенно ответить не может. Однако дело в том, что в некоторых случаях, кажется, всё-таки может.
Речь идет о тех случаях, когда биологи находят, будто бы, пути эволюции самих белков - когда белок сначала, вроде бы, выполнял одни функции, а потом, после некоторой генетической модификации, стал выполнять другие функции.
Например, мы видим, что аминокислотная последовательность в каком-то белке, который выполняет, допустим, функцию «переваривания пищи» – очень похожа на аминокислотную последовательность другого белка, но который занимается уже совершенно другим делом. Например, предохраняет организм от переохлаждения - от образования льда внутри тканей организма.
Это я сейчас озвучиваю получивший широкое распространение пример с так называемыми белками «антифризами» у полярных рыб. Понятно, что этот факт был открыт в ходе узко специфичных биологических исследований. Однако дарвинисты-миссионеры постарались сделать эту информацию широко известной. Логика их вполне понятна – ведь этот факт, вроде бы, куда больше свидетельствует в пользу случайной эволюции (методом случайных мутаций), чем в пользу разумного планирования.
Действительно, белки «антифризы» некоторых антарктических рыб имеют сходство аминокислотной последовательности с другим белком - панкреатической трипсиногеноподобной протеазой, который занимается совершенно другой деятельностью - «перевариванием пищи» (расщеплением пептидной связи). С точки зрения разумного планирования, переделывать имеющийся белок в какой-то другой, чтобы он выполнял совершенно другую функцию, кажется, не слишком логично. Правильнее, наверное, было бы создать отдельный белок, который бы сразу (исходно) выполнял возложенную на него специальную задачу. Но нет. Мы ясно видим именно следы «переделки». В этом переделанном белке сохранились даже некоторые участки от прежнего белка, которые, вроде бы, не нужны новому белку для выполнения его работы. А достались ему просто «в наследство» от исходного белка.
То есть, здесь мы, кажется, имеем такую ситуацию, про которую было хорошо спето в одной популярной песенке:
- Я его слепила из того, что было, а потом что было, то и полюбила (С)
Понятно, что такое положение дел больше всего свидетельствует в пользу версии случайной эволюции. Действительно, именно от случайной эволюции, прежде всего, можно ожидать «творений» по принципу «слепила из того, что было». Ведь естественная эволюция слепа, а выживать рыбам в антарктических водах как-то было надо. Особенно если предположить, что эти антарктические воды несколько миллионов лет назад начали постепенно охлаждаться. Вот слепая эволюция в этих условиях и стала подхватывать такие аминокислотные изменения в любых случайных белках, которые позволяли этим белкам «прилипать» к нарождающимся кристалликам льда, не давая этим кристалликам разрастаться дальше и разрушать живые ткани. И таких подхваченных случайных решений оказалось довольно много. Например, белки-антифризы из группы АФГП были обнаружены у антарктических рыб семейства нототениевые (Nototheniidae) и как уже говорилось, предположительно являются эволюционной «переделкой» какой-то трипсиногеноподобной протеазы. Причем у одного из видов нототениевых рыб (антарктического клыкача) найден химерный ген с промежуточными свойствами, кодирующий как АФГП полипротеин, так и протеазу (Бильданова и др., 2012). То есть, найдена как бы переходная форма между белком трипсиногеном и белком-антифризом.
А вот белки-антифризы группы АФП I (характерные уже для других рыб) похожи на продукт переделки уже других белков – установлена гомология между геном АФП I типа рыб и генами белков хориона и кератина липариса атлантического (Бильданова и др., 2012). А белки-антифризы группы АФП II, возможно, произошли от какого-то лектиноподобного предкового белка (Бильданова и др., 2012). Наконец, белки-антифризы группы АФП III (имеющиеся у антарктических бельдюговых рыб) произошли, скорее всего, от древнего гена синтазы сиаловой кислоты, путем дивергенции и дупликации (Бильданова и др., 2012).
Как видим, решений получилось много. И все эти решения в итоге позволили разным рыбам выживать в очень холодных арктических и антарктических водах.
Но и это еще не всё. Разнообразные белки «антифризы» были обнаружены еще и у насекомых (причем в разных таксонах насекомых - разные белки-антифризы). Еще у антарктической нематоды (круглый червь), у грибов, у диатомовых водорослей, в разных таксонах растений (разные белки антифризы), у бактерий (Бильданова и др., 2012).
В общем, такое ощущение, что молекулярная филогения именно белков-антифризов четко вписывается в сценарий естественной эволюции (т.е. дарвинизма). Например (рисуем возможный сценарий), антарктические воды стали постепенно охлаждаться, и сразу у разных групп антарктических рыб естественный отбор начал отыскивать разные мутации в разных белках, лишь бы эти изменения подходили для выполнения «антифризной» роли. И естественный отбор находил то, что ему было нужно - отдельные особи, у которых происходили такие мутации (неважно в каких генах), где аминокислотная последовательность мутантного белка начинала «липнуть» к кристалликам льда (препятствуя их объединению и росту), получали преимущество в выживании. И в конечном итоге получилось то, что получилось – во-первых, разнообразие антифризных белков у разных групп арктических и антарктических рыб, а во-вторых, явно «переделочный» характер этих белков.
Причем такую же картину мы видим и по белкам-антифризам в других биологических таксонах – у насекомых, или, например, у растений – белки-антифризы тоже отличаются разнообразием и нередко демонстрируют гомологию (сходство) с какими-то другими белками, выполняющими какие-то совершенно другие функции.
В общем (еще раз) создаётся такое ощущение, что здесь всё прямо по Дарвину. Так неужели это как раз тот случай, когда факты молекулярной филогенетики говорят (свидетельствуют) именно в пользу дарвиновских механизмов эволюции?
Хотя окончательным доказательством такие факты всё-таки не являются. Потому что это всё же мог быть разумный дизайн. Но тогда это был весьма специфичный разумный дизайн. Который, с одной стороны, можно было бы назвать в высшей степени «ленивым». Ведь этот дизайн был выполнен по принципу «из того, что было». И еще, кажется, по принципу «тяп-ляп и готово».
С другой стороны, «ленивым» этот дизайн тоже никак нельзя назвать. Ведь хотя белок-антифриз создаётся как бы «из того, что было», но зато у разных рыб это «из того что было» каждый раз оказывается разным. А ведь если бы дизайнер был ленив, то он бы, наверное, поленился переделывать каждый раз разные белки (превращая их в разные белки-антифризы), а воспользовался бы каким-нибудь единственным (единожды созданным) белком-антифризом, снабдив им (и только им) все имеющиеся виды рыб, которые сталкиваются с проблемой переохлаждения. Однако такое ощущение, что гипотетический дизайнер просто забавлялся, «вылепляя» белки антифризы каждый раз из разных (имевшихся под рукой) белков независимо для каждой отдельной группы рыб. Такое поведение уже трудно назвать «ленивым». В общем, получается какая-то ерунда - наш воображаемый дизайнер получается как бы и ленив и не ленив одновременно.
Тем не менее, даже такие причуды, в принципе, дизайнеру делать не запретишь. Ведь на то он и дизайнер, чтобы поступать, как хочет.
Кроме того, можно предположить и некоторые другие гипотезы, связанные с дизайном. Например, дизайнеров могло быть много. И каждый дизайнер занимался проектом какого-то своего (конкретного) таксона рыб, особо не вникая в то, что там творили его «соседи-дизайнеры» с другими таксонами рыб.
Еще можно предположить, что дизайнеры пользовались «языком программирования» очень высокого уровня, где даются лишь общие указания, «что нужно сделать», а не «как это сделать». Программисты поймут, что я имею в виду, потому что именно такой принцип используется сегодня в некоторых языках программирования сверхвысокого уровня. То есть, дизайнеры могли давать своим «программам редактирования живых существ» лишь самые общие указания – например: «Пусть эта рыба станет жить в холодной воде». И далее «программа редактирования живых существ» уже сама реализовывала эту инструкцию тем или иным способом (может быть, не самым мудрым из всех возможных).
Наконец, можно предположить, что дизайнер вообще не «заморачивался» с каждым конкретным белком и каждой конкретной рыбой, а просто встроил некий «решатель проблем» в геномы самих рыб. И в результате сами рыбы оказались способны решать некоторые простые вопросы своего выживания – допустим, самостоятельно приспосабливаться к охлаждению воды, или, наоборот, к её потеплению. Например, позвоночные животные имеют аналогичный «встроенный решатель проблем» в своей иммунной системе. Когда организм позвоночного животного сталкивается с инфекцией, в клетках его иммунной системы происходит быстрый подбор нужного антитела к этой инфекции. За счет того, что имеющаяся (от рождения) «база данных» антител подвергается очень быстрому перебору и модификации с целью поиска нового варианта антитела, подходящего конкретно в данной ситуации. Это осуществляют специальные ферменты, обеспечивающие гипер-мутирование на нужных участках ДНК. В результате, организм подбирает нужный «ключ» к напавшей на него инфекционной болезни. Это весьма эффективный механизм иммунитета, хотя, конечно, возможности его тоже не безграничны. Примерно так же может быть и в разбираемом нами случае – в принципе, рыбы могут иметь какие-то встроенные в их геномы генетические подпрограммы, специально предназначенные для решения некоторых частных проблем их выживания. Хотя возможности этих генетических подпрограмм тоже не беспредельны.
Я не знаю, убедительно ли прозвучали для читателя только что перечисленные гипотезы с участием разумного дизайна в появлении белков-антифризов. Лично мне эти гипотезы самому показались неубедительными. Создаётся ощущение, что озвученные варианты «притянуты за уши», лишь бы только в них всё-таки фигурировал разумный дизайн (а не естественный отбор).
Гораздо проще согласиться, что белки-антифризы полярных рыб – больше всего свидетельствуют именно в пользу естественной эволюции по дарвиновским механизмам. Потому что именно от естественной эволюции по (слепым) дарвиновским механизмам, прежде всего, ожидается именно такая картина - разные белки-антифризы в разных группах рыб, полученные в результате переделки других белков. Всё это очень похоже именно на результат независимых естественных эволюционных процессов (на молекулярно-генетическом уровне) под давлением одних и тех же факторов среды и соответствующего естественного отбора в этой среде.
Тем не менее, озвученные выше версии разумного дизайна, хотя и выглядят откровенными спекуляциями, теоретически всё же не исключены. Поэтому после долгих колебаний я всё-таки решился почитать про эти белки-«антифризы» более подробно. Дело в том, что, не являясь генетиком, я обычно стараюсь избегать погружения в такие вопросы, и поэтому долго не решался на чтение оригинальных публикаций в этой области. Но наконец решил, что это сделать всё-таки надо.
И сразу же выяснились чрезвычайно интересные вещи.
Итак, открываем соответствующий обзор и начинаем читать (Бильданова и др., 2012:250):
Первые АФП описаны более 40 лет назад (DeVries, Wohlschlag, 1969).[49] С тех пор выявлено множество их типов у представителей разных таксонов (табл. 1) (Jia, Davies, 2002). Судя по вариабельности связывающихся со льдом белков и их неупорядоченному филогенетическому распространению, эти белки возникали независимо, в основном как инструмент приспособления вида к новой экологической нише или в связи с изменениями климата (Cheng, 1998; Doucet et al., 2009; Deng et al., 2010).
Пока всё вроде бы выглядит так, как мы и писали выше – «вариабильность» и «неупорядоченное филогенетическое распространение» этих белков свидетельствует в пользу их независимого возникновения в ходе (видимо) случайной эволюции. То есть, начинается, вроде бы, «за здравие». Однако посмотрим, чем всё закончится.
В разделе «Происхождение и эволюция АФП» читаем (Бильданова и др., 2012:262):
Группа АФП характеризуется недавним эволюционным происхождением, множественностью компонентов у одного организма, независимым происхождением у представителей близкородственных таксонов и конвергентным сходством у неродственных групп организмов (Barrett, 2001)…
Вот как только я прочитал фразу про «конвергентное сходство», так сразу же и насторожился. Потому что «конвергентное сходство» - это, на самом деле, такая штука, которая на дарвинистском языке означает явление, не слишком хорошо объяснимое в рамках дарвинизма, но зато прекрасно объясняемое в рамках разумного дизайна.
И предчувствия нас не обманули. Начинаем читать про антифризные белки первой группы, и выясняем, что белки этой группы обнаружены у антарктических нототениевых рыб… но не только у них! Сходные антифризные белки обнаружены еще и у сайки (Boreogadus saida). А ведь сайка (полярная тресочка) принадлежит вообще к другому семейству (тресковые) и даже к другому отряду рыб (трескообразные). Более того, сайка вообще не пересекается с представителями семейства нототениевые еще и географически, поскольку плавает не в холодных антарктических водах южного полушария (где обитают нототениевые рыбы), а населяет столь же холодные арктические воды северного полушария. То есть, обитает, в буквальном смысле, на другом конце Земного шара.
Получается, что « почти идентичные АФГП» (Бильданова и др., 2012:262) имеются у разных (независимых) семейств рыб (нототениевых и тресковых), которые еще и обитают на противоположных концах Земли.
Как это понимать? Почему наша «случайная эволюция», о которой мы только что говорили, как о наиболее правдоподобном объяснении «эволюции белков-антифризов», совершила столь странное «совпадение»? Где, собственно, заявленная случайность этой эволюции? Мы только что, вроде бы, пришли к выводу, что белки-антифризы лепились из других белков по случайному принципу (помните: «я его слепила из того, что было»), как средство «экстренной эволюционной помощи при похолодании или смене экологической ниши». Но когда мы перешли к изучению конкретных фактов, неожиданно оказывается, что эта «лепка» совсем не выглядит случайной - она была повторена в совершенно независимых таксонах рыб.
Но ведь мы только что озвучили практически то же самое в версии про «ленивого дизайнера». Помните? Мы предположили, что «ленивый дизайнер» вылепил бы нужный белок из первого же более-менее подходящего (для этой цели) другого белка… а потом бы распространил такое же решение на другие таксоны рыб, которые сталкиваются с такими же экологическими проблемами. Но мы отбросили эту версию, как «притянутую за уши». Помните?
И вот оказывается, что необходимый АФГП был «вылеплен» из другого белка, и задействован в двух совершенно разных таксонах рыб, которые сталкиваются с одинаковыми экологическими проблемами. То есть, наше рассуждение про «ленивого дизайнера»… неожиданно подтвердилось с точностью до буквы.
Вот так неожиданно ситуация развернулась ровно на 180 градусов. Буквально только что (пока мы не вдавались в подробности) эта ситуация выглядела совершенно как «эволюция случайных генов под действием сходных условий среды». А разумный дизайн плёлся где-то на задворках в качестве «притянутой за уши невероятной спекуляции». Но стоило нам ознакомиться с конкретными фактами, как теперь нам уже необходимо защищать нашу теорию о «случайной эволюции из случайных генов», изобретая дополнительные объяснения, почему это наша «случайная эволюция» демонстрирует столь серьезные повторы своих «случайных решений».
И объяснения этому факту, конечно, приводятся (в рамках дарвинизма). Считается, что это была действительно независимая эволюция, которая, тем не менее, привела к конвергентному (генетическому) сходству. На независимый характер этой эволюции указывает то, что несмотря на общее большое сходство, между АФГП нототениевых и тресковых имеется и целый ряд отличий (Бильданова и др., 2012:262):
…гены АФГП нототениевых и тресковых имеют отличия, которые убедительно показывают независимую эволюцию АФГП-генов трески: а) различные сигнальные пептидные последовательности; б) разные последовательности спейсеров, соединяющих отдельные гены АФГП в полипротеине, что приводит к различным механизмам процессинга предшественников полипротеина; в) разные последовательности, кодирующие повторы Thr-Ala/Pro-Ala; г) геномные локусы АФГП гена трески и нототениевых АФГП тоже отличаются (Cheng, 1998)…
В результате авторы обзора приходят к выводу:
Таким образом, почти идентичные АФГП двух не связанных групп рыб представляют собой пример конвергенции белковых последовательностей, т.е. развитие аналогичного белка из разных предков в результате воздействия одинаковых условий окружающей среды.
Ну что же. Эволюционное объяснение найдено. Оказывается, почти идентичные белки вполне могут возникать из разных предковых белков, если того требуют одинаковые условия среды. Действительно, можно предположить, что функция связывания со льдом требовала примерно сходных аминокислотных последовательностей в белке. Вот в результате этих требований (и соответствующей эволюции под действием естественного отбора) и получилось итоговое сходство обсуждаемых белков.
Правда, мы совсем недавно писали, что имеется большое разнообразие белков-антифризов. Поэтому, казалось бы, АФГП тресковых и нототениевых было бы совсем не обязательно становиться почти идентичными в ходе «случайной эволюции». Но вот теперь, оказывается, что обязательно. Ну что же, простим этот маленький каприз нашей «случайной эволюции». Ведь, как известно, неисповедимы пути её.
Тем не менее, еще раз отметим, что версия «случайной эволюции» становится уже далеко не такой «очевидной», какой она выглядела до ознакомления с конкретными антифризами тресковых и нототениевых рыб. В то же время такие (почти идентичные) решения, применённые в разных (независимых) биологических таксонах - уже гораздо лучше вписываются в концепцию разумного дизайна этих таксонов, чем это выглядело вначале.[50]
Но может быть, «почти идентичность» АФГП тресковых и нототениевых – это просто такое случайное исключение из правил? Может быть, АФП других организмов ясно демонстрируют совершенно случайный характер своего образования?
Ну что же, давайте посмотрим. Читаем про антифризные белки следующей группы (Бильданова и др., 2012:262):
АФП I типа рыб являются полифилетическими по происхождению, они обнаружены у представителей трех неродственных отрядов костистых рыб и представляют собой пример конвергентной эволюции (Hobbs et al., 2011).
Вот это да! Оказывается, удивительная ситуация со странными повторами… повторяется снова. Причем теперь уже в более тяжелом варианте. В данном случае получается, что антифризные белки АФП I имеются уже у трех неродственных отрядов рыб. Конкретно в работе озвучивается керчак (скорпенообразные), зимняя камбала (камбалообразные) и губан (окунеобразные) (Бильданова и др., 2012:252).
Как объяснить теперь уже эти независимые повторы? Если это не «разумный дизайн» (сходные инженерные решения, примененные для выполнения сходных задач в совершенно разных биологических таксонах), то тогда что же это такое?
В рамках эволюционных воззрений – это тройное (!) совпадение опять «списывается» на конвергенцию. То есть, антифризные белки этой группы так похожи друг на друга у представителей трех разных отрядов рыб просто потому, что… так получилось. Может быть, на роль будущих антифризных белков были избраны похожие исходные белки (чисто случайно, конечно). Например, вышеупомянутые белки хориона или кератина во всех трех отрядах рыб (просто случайно так совпало). Или же исходные белки были разные, но одинаковые требования к этим белкам (реагировать с кристаллами льда) в итоге привели к сходной аминокислотной последовательности в исходно разных белках. Несмотря на то, что те же самые требования к белкам привели (почему-то) к другой аминокислотной последовательности у тресковых и нототениевых. А у сельди и корюшки – к третьей аминокислотной последовательности (см. ниже). И у диатомовых водорослей – тоже к другой. У пшеницы – еще к другой. У моркови – тоже к другой. У грибов – опять к другой. У бабочек – снова к другой. У жуков – снова к другой. И у бактерий… правильно, тоже к другой. Итак, у всех вышеперечисленных организмов, «одинаковые требования среды» привели к другим антифризным белкам. А вот в трех независимых отрядах рыб (скорпенообразных, камбалообразных и губановых) эти же требования среды почему-то потребовали от аминокислот выстроиться именно в определенные (сходные) аминокислотные последовательности. Которые сами же дарвинисты непременно объявили бы гомологичными (т.е. происходящими от одного предка), если бы эти белки не были найдены в трех независимых таксонах рыб.
Да, именно так нас и учит теория эволюции (даёт вот такие объяснения). В одних случаях случайная эволюция совершенно свободно «вылепляет» самые разные антифризные аминокислотные последовательности из самых разных белков. И у неё всё получается. А в других случаях эта же случайная эволюция не может поступить никак иначе, кроме как повторить сходную антифризную аминокислотную последовательность три независимых раза подряд.
Но может быть, это совсем не совпадение? Может быть, для такого решения имелись какие-то резонные основания? И именно поэтому наша «случайная эволюция» и прошла по одному и тому же пути три раза подряд? Но если мы сделаем такое предположение, то сразу же поймем, что точно такое же предположение можно сделать и в рамках концепции разумного дизайна. Что имелись какие-то (резонные) основания создать именно такую (а не какую-то другую) антифризную аминокислотную последовательность в этих трех конкретных случаях. И именно поэтому некий разумный дизайнер так и сделал – применил сходное решение три раза подряд в тех независимых отрядах рыб, которым это решение подходило (по каким-то причинам).
Однако мы пока делаем вид, что как бы верим эволюционным объяснениям (про многочисленные «конвергенции»). Действительно, пока еще ничего особо невозможного не было озвучено. Ну, совпало так три раза подряд, что «мишенями» для случайных мутаций при создании белков-антифризов в трех разных отрядах рыб послужили какие-то, допустим, исходно похожие белки. Поэтому продолжаем читать соответствующий обзор.
Переходим к чтению про белки-антифризы следующей группы… И теперь уже падаем со стула. Читаем (Бильданова и др., 2012:263):
Независимое появление АФП II типа в трех неродственных ветвях костистых рыб (сельди, корюшки и морского ворона) является экстраординарным по нескольким причинам (Graham et al., 2008a). Эти гомологи лектинов являются единственными лектинами, у которых пятый дисульфидный мостик специфически локализован, и они гораздо более похожи друг на друга, чем на любой другой гомолог лектинов. Их сходство распространяется и на уровень последовательности ДНК, где даже интроны имеют до 97% гомологии. В базах данных не были обнаружены родственные аминокислотные последовательности других видов с гомологией более 40%. Это согласуется с данными геномной Саузерн гибридизации, которая показала отсутствие гомологов АФП второго типа у других видов рыб. Консервативность интронов и экзонов, отсутствие корреляции между эволюционным расстоянием и скоростью мутаций, соотношение синонимичных и несинонимичных замен в кодонах не соответствуют гипотезе о существовании предшественника этих генов, который был элиминирован во всех таксонах костистых рыб.
Некоторые авторы считают, что горизонтальный перенос гена АФП II типа является наиболее вероятным объяснением большой гомологии аминокислотной последовательности (до 85%), низкой частоты синонимичных мутаций и высокой консервативности числа, положения и последовательности интронов (Graham et al., 2008a).
То есть, ситуация становится уже просто скандальной. АФП II типа точно так же как и предыдущие (АФП I типа) обнаружены у представителей сразу трех неродственных отрядов рыб (сельдеобразные, корюшкообразные и скорпенообразные). Причем сельдеобразные принадлежат даже к другому надотряду (Сlupeomorpha).
Между тем, уровень гомологии аминокислотной последовательности доходит до 85%. И это сходство распространяется даже на интроны!
Понятно, что лучше всего наличие столь сходных генов в разных таксонах рыб объясняется в рамках разумного дизайна. Как я уже говорил, для разумного дизайна совершенно естественно, что одинаковые инженерные решения (одинаковые гены) используются в разных таксонах рыб для решения одних и тех же задач.
А вот в рамках теории естественной эволюции, очевидно, что такое сильное сходство белков и генов уже нельзя списать ни на какую «конвергенцию». Потому что такой уровень сходства уже слишком невероятен для любых случайных совпадений.
И вот нашей многострадальной «теории эволюции» в данном случае приходится «доставать из широких штанин» (С) уже другую дополнительную (ad-hoc) гипотезу для объяснения еще и этого факта сходства белков и генов в трех совершенно неродственных таксонах. Как видно из приведенной цитаты, некоторые авторы предлагают применить здесь объяснение «горизонтальным переносом». Это значит, что нам предлагают считать следующее – данные рыбы просто «нахватались» друг у друга соответствующих генов. Правда, нахватались они этих генов таким способом, который пока неизвестен современной науке. Ведь науке пока известен только один способ «пересадки» генов от одних эукариотических организмов к другим – это методы генной инженерии. Однако рыбы перечисленных выше отрядов (сельдеобразные, корюшкообразные и скорпенообразные) вряд ли владеют методиками генной инженерии. У этих рыб нет соответствующего оборудования. И даже нет рук, чтобы на этом оборудовании работать.
Поэтому считается (теоретически), что случайную пересадку гена мог бы произвести какой-нибудь вирус, который, во-первых, случайно «выхватит» нужный ген из ДНК организма-хозяина и случайно встроит в свою ДНК. После этого (во-вторых), этот вирус должен случайно заразить не того хозяина, к которому он приспособлен (например, атлантическую сельдь), а какую-нибудь совершенно постороннюю рыбу, например, азиатскую корюшку. В-третьих, этот вирус должен заразить не абы какую клетку нового организма-хозяина, а именно его половую клетку. Ведь если вирус встроит своё ДНК в клетку какой-нибудь мышцы, или в клетку крови, или печени, то пересаживаемый ген там так и останется, и умрет вместе с данной рыбой. И наконец, встроенный ген (после столь удачного попадания в нужное место), должен каким-то образом еще и начать там удачно работать. То есть, когда он начнет работать, это должно оказаться полезным (а не вредным) для организма. Поэтому он должен начать работать именно там где нужно, и в то время, когда это нужно. Ведь что получится, если этот ген «врубится» в тот момент, когда, допустим, рыбий эмбрион только начал формировать себе мозг… И в это время включается ген, штампующий в этом мозгу белки-антифризы… Что из этого получится, знает только один Ричард Докинз. Наверное, данная рыба сразу же перейдет на совершенно новый этап эволюции.
Естественно, воочию таких чудес («горизонтальных переносов у эукариот») не видел еще ни один биолог. Но поскольку «чужие» гены обнаруживаются (в самых разных организмах) всё чаще и чаще, то биологам и приходится теоретически допускать возможность таких вот чудес. Действительно, не допускать же в качестве объяснения разумный дизайн. Ведь это так невероятно - редактирование генетических программ методами генной инженерии. Несмотря на то, что мы (люди) уже сами занимаемся подобной деятельностью (спустя всего 60 лет после того, как разобрались, что такое ДНК). Но допускать, что подобной деятельностью мог бы заниматься кто-нибудь еще (кроме нас), мы не хотим ну прямо вот никак.
В общем, давайте подводить итоги.
Сначала вспомним, как прекрасно всё начиналось – при взгляде на белки-антифризы «с высоты птичьего полёта» наша любимая теория естественной эволюции так хорошо подтверждалась… А несчастный разумный дизайн выглядел так наивно.
А теперь давайте посмотрим, к чему мы пришли в итоге:
1. Совпадение АФГП у нототениевых и тресковых рыб – выглядит, конечно, похожим на разумный дизайн, но это не разумный дизайн, а просто «результат конвергенции в ходе случайной эволюции».
2. Совпадение АФП первого типа у скорпенообразных, камбалообразных и окунеобразных – это, конечно, тоже похоже на разумный дизайн. Но мы не будем «верить глазам своим», а будем тоже считать это «результатом конвергентной эволюции».
3. Ну а совпадение АФП второго типа у сельдеобразных, корюшкообразных и скорпенообразных – это уже вообще, настолько похоже на разумный дизайн, что, собственно, и не отличить. Это явные следы генной инженерии. Однако мы и это тоже не будем считать разумным дизайном. А объясним теперь уже «горизонтальным переносом» соответствующих генов. Потому что предыдущее объяснение про «результат конвергентной эволюции» тут уже явно не «катит».
Итак, что мы наблюдаем в итоге? В итоге мы наблюдаем, что начали мы «за здравие», да вот только закончили «за упокой». Вначале у нас нарисовалась такая красивая картинка по эволюции белков-антифризов, причем именно методом естественного отбора случайных мутаций (у случайных белков). Однако эта благостная картинка буквально рассыпалась в прах, стоило ей соприкоснуться с конкретными фактами. Получилось, что реальные установленные факты по белкам-антифризам не столько «подтверждают эволюцию», сколько задают загадки и создают парадоксальные ситуации.
Причем то же самое распространяется не только на рыб, но и на другие таксоны, и даже на целые царства живых существ. Читаем (Бильданова и др., 2012:263):
В последнее время были клонированы АФП диатомовых водорослей (Janech et al., 2006) и антарктической бактерии (Raymond et al., 2007), эти белки показали высокую гомологию с АФП грибов (Xiao et al., 2010). Есть все основания считать, что гомологи АФП грибов широко распространены в нескольких царствах, благодаря возможному горизонтальному переносу генов между эукариотическими микробами и прокариотами (Raymond et al., 2007).
Как говорится, «совсем приехали». Теперь уже АФП совпадают у диатомей, бактерий и грибов. Что заставляет привлекать палочку-выручалочку с «горизонтальным переносом» еще и для этих таксонов. То есть, на самом деле, столь разрекламированные (некоторыми проповедующими дарвинистами) белки-антифризы не только не «подтверждают эволюцию», а наоборот – эту бедную «теорию эволюции» приходится буквально спасать от тех фактов, которые открылись при изучении белков-антифризов. Спасать при помощи дополнительных гипотез про «горизонтальные переносы» между разными царствами живых существ. Причем мы видим, что для спасения теории эволюции (от наступающих на неё фактов) применение гипотезы «горизонтального переноса» должно иметь массированный характер.
Но и это еще не всё. Помните, рассуждая выше о том, как мог бы осуществляться разумный дизайн белков-антифризов, я высказал (в том числе) мысль о возможном наличии некоего готового «решателя проблем», встроенного в геномы соответствующих животных. Вспомнили?
А теперь давайте прочитаем следующую цитату (Бильданова и др., 2012:263):
АФП III и АФГП отличаются по составу и структуре, они выявлены в различных систематических таксонах, однако сходная структура генов этих АФП свидетельствует о наличии у рыб общего механизма организации участков геномов, отвечающих за приспособление к экстремальным условиям полярных ареалов (Deng et al., 2010).
То есть, это либо свидетельство прямого разумного дизайна, проявившее себя в организации сходной структуры генов при решении сходных «технических» задач в разных биологических таксонах. Либо у рыб имеется встроенный «автоматический решатель» подобных экологических проблем (тоже, видимо, «заботливо предоставленный производителем»).
Да и вообще. Сегодня уже понятно, что многочисленные парадоксальные ситуации складываются постоянно не только при попытках проследить пути генетической эволюции рыб (или их белков), но и в любых других биологических таксонах. Это и есть та самая суровая реальность, которую выше я упомянул, но пока обещал игнорировать. А может быть, хватит уже игнорировать?
Ведь из простого перечисления установленных (к сегодняшнему дню) генетических конфликтов можно уже, наверное, составить целую книгу.
Прежде всего, это будет длинный перечень генетических фактов, которые резко конфликтуют с морфологическими фактами. Еще хуже получается, когда генетические исследования, проведенные по одним генам, вступают в конфликт с результатами таких же генетических исследований, но проведенных по другим генам этих же самых (!) организмов. То есть, генетические факты («доказывающие эволюцию»?) на самом деле, весьма часто противоречат: 1) друг другу, 2) общепринятым сценариям эволюционного происхождения биологических таксонов.[51]
Для любопытствующих приведу типичный пример - резюме очередной научно-популярной заметки, с говорящим названием «Игуаны голосуют против молекулярной филогенетики» (Наймарк, 2012):
Одновременно опубликованы две работы, касающиеся филогении чешуйчатых (ящериц, змей, амфисбен и их родичей). Одна из них реконструирует молекулярную эволюцию ядерных генов, вторая — сравнивает морфологические признаки вымерших и ныне живущих представителей. Два филогенетических дерева оказались принципиально несхожи. В особенности это касается игуан, имеющих множество примитивных черт, но оказавшихся на молекулярном дереве среди своих продвинутых четвероюродных кузин. Этот методологический конфликт пока не удается разрешить.
Это пример филогенетического конфликта на уровне разных биологических семейств. Но еще более интересны такие генетические конфликты, которые обнаруживаются на уровне биологических родов, и даже биологических видов. Такие примеры интересны потому, что на самых «мелких» уровнях таксономической иерархии эволюционные связи между живыми существами, казалось бы, должны быть видны наиболее отчетливо.
Действительно, если мы сравниваем между собой таксоны, далекие друг от друга, то здесь филогенетические взаимоотношения как бы «размываются». Становясь тем более неопределенными, чем выше ранг этих биологических таксонов. Ведь далекие биологические группы серьезно отличаются друг от друга по целому комплексу признаков (морфологических, анатомических, физиологических, экологических, биохимических). Одновременно, гены этих (далеких друг от друга) биологических групп тоже демонстрируют весьма серьезные различия в своих нуклеотидных последовательностях. В значительном числе случаев «родство» этих генов угадывается лишь на уровне самых консервативных отдельных участков. В то время как остальная (наибольшая) часть их нуклеотидных последовательностей – вообще не имеют между собой ничего общего. В этом случае становится не слишком понятно, считать ли это результатом глобального расхождения нуклеотидных последовательностей (от исходных генов «общего предка») в ходе очень долгой эволюции? Или же так и было задумано сразу - чтобы эти белки были разными у столь разных организмов? То есть, сила «генетических доказательств эволюции» в случае далеких биологических таксонов серьезно снижается (во всяком случае, в глазах критиков).
И совсем другое дело, когда мы наблюдаем весьма близкие нуклеотидные последовательности в близких биологических таксонах. Например, у разных биологических видов в рамках одного и того же рода. В этом случае становится, действительно, непонятно, зачем гены, почти идентичные друг другу и выполняющие у этих близких видов одинаковые функции, тем не менее, всё-таки немного различаются? Напрашивается мысль, что эти различия (между почти одинаковыми генами) не имеют никакого биологического смысла, а были приобретены просто случайно, в результате мутаций, накопленных за определенный период времени.
Но ведь мы наблюдаем, что между разными биологическими видами в пределах одного и того же рода - гены различаются меньше, чем между видами, принадлежащими к разным родам (и тем более, к разным семействам). Именно этот факт, «помноженный» на постулат о чисто случайном («накопительном») характере подобных различий - и приводит нас к выводу об имевшей место эволюции. Где разные биологические виды в рамках одного и того же рода имеют меньше отличий (в почти одинаковых генах) потому, что эти биологические виды разделились от общего эволюционного предка сравнительно недавно (и поэтому успели накопить меньше различий в генах). А биологические виды из разных биологических родов имеют больше отличий (в сходных генах) потому, что они отделились от общего эволюционного предка раньше (и поэтому успели накопить больше различий в соответствующих генах).
Таким образом (еще раз) свидетельства эволюции, «добытые» из геномов близких биологических видов и родов, казалось бы, должны быть наиболее яркими. Тем не менее, филогенетические конфликты встречаются даже на этом уровне. Причем регулярно.
В качестве «легкой разминки», приведу двух разных сомиков из рода Prietella. На этот пример я натолкнулся чисто случайно, когда знакомился с таксономическим разнообразием пещерных животных.
Изучая видовой состав пещерных рыб Мексики, я узнал о существовании двух видов слепых мексиканских сомиков - Prietella phreatophila и Prietella lundbergi. Эти два пещерных создания по комплексу морфологических признаков весьма близки друг к другу. Понятно, что значительная часть их сходных признаков связана с пещерным образом жизни (Walsh & Gilbert, 1995).[52] Однако, помимо чисто «пещерных» признаков, у обоих видов сомиков Prietella имеется и ряд специфических общих морфологических черт, которые не имеют прямого отношения к обитанию в пещерах. Эти специфические признаки, характерные для обоих видов рода Prietella, отсутствуют у представителей других родов этого семейства (в том числе и у других пещерных видов из этого же семейства).
Очевидно, что такие признаки могли появиться (сразу у обоих видов рода Prietella) либо в результате сходного дизайна (в рамках концепции разумного дизайна), либо в результате эволюции от общего предка – просто в качестве морфологического наследия, доставшегося им от этого предка.
Поэтому авторы соответствующей работы пришли к выводу, что род сомиков Prietella имеет монофилетическое происхождение. То есть, оба вида этого рода происходят от общего эволюционного предка (Walsh & Gilbert, 1995).
Однако такой эволюционный сценарий, построенный на совершенно очевидной морфологической близости двух видов сомиков… подвергся неожиданной атаке со стороны генетических фактов. В более поздней работе (Wilcox et al., 2004) авторы исследовали митохондриальную ДНК двух разных видов рода Prietella. И оказалось, что два обсуждаемых сомика по митохондриальной ДНК – не родственны друг другу. То есть, не имеют общего предка. Один из видов сомиков (P. phreatophila) оказался генетически ближе к роду Ameiurus (другому роду сомов из этого же семейства Ictaluridae, кошачьи сомы). А второй слепой пещерный сомик, Prietella lundbergi, оказался генетически ближе уже к роду Ictalurus (тоже из этого же семейства).
Более того, авторы предыдущей работы (Walsh & Gilbert, 1995) по совокупности морфологических признаков сделали вывод о родстве всего рода Prietella к роду Noturus (еще одному роду из этого семейства). А вот генетические исследования озвучили уже другие родственные роды: Ameiurus и Ictalurus (Wilcox et al., 2004).
Интересно, что авторы генетического исследования (Wilcox et al., 2004) признают сильную морфологическую близость видов P. phreatophila и P. lundbergi. Но предлагают считать эту близость результатом конвергенции. То есть, результатом независимой эволюции, которая, тем не менее, привела к очень сходным биологическим формам.
Однако конвергенцию (независимое приобретение морфологического сходства) в рамках дарвинизма объясняют давлением одинаковых условий среды. Но в нашем случае так можно объяснить только часть общих признаков обсуждаемых сомов. А с другими морфологическими признаками, которые у этих видов тоже общие, но не связаны с пещерными условиями, так и остаётся непонятно, что делать.[53]
В своё оправдание авторы «генетической» работы отмечают (Wilcox et al., 2004):
«Независимое происхождение P. lundbergi и P. phreatophila удивительно на основе морфологии, но зато имеет значительный смысл в отношении биогеографических перспектив»
Здесь авторы работы намекают еще на одно интересное обстоятельство. Дело в том, что ареалы этих двух пещерных сомиков – географически разнесены друг от друга на очень значительное (по меркам пещерных видов) расстояние. Между крайними точками этих ареалов пролегает, как минимум, 600 км засушливой территории. Причем подземные ареалы этих сомиков относятся к бассейнам разных рек, которые разделены между собой горным массивом (Walsh & Gilbert, 1995). Поэтому представить себе, каким образом эти виды пещерных сомиков (или общий пещерный предок этих сомиков) попали в те подземные комплексы, в которых они обитают сейчас – очень трудно. Этот факт представляет собой определенную биогеографическую загадку, над которой раздумывают авторы сразу трех работ, посвященных этим рыбкам (Walsh & Gilbert, 1995; Hendrickson et al., 2001; Wilcox et al., 2004).
Вот поэтому авторы работы (Wilcox et al., 2004) и пишут (см. цитату выше) о том, что генетические факты противоречат морфологическим фактам, но зато становится менее загадочным географическое распространение этих видов рыб (поскольку генетические факты указывают на их независимое происхождение).
Таким образом, здесь переплетаются в узел разные группы фактов, конфликтующие между собой – морфологические, генетические и биогеографические.
Но зачем мы сейчас копаемся в таких мелочах, как генетические и биогеографические странности двух несчастных видов сомиков? В конце концов, не все ли равно, происходят эти сомики от общего предка (на что указывает их морфология) или от разных предков, относившихся к разным родам этого же семейства (как показало генетическое исследование).
Еще раз повторюсь – нет, такие примеры, наоборот, очень интересны. Например, свидетели Дарвина постоянно рассказывают о том, что в результате генетических исследований, наиболее близкими к роду человек (Homo) в генетическом плане оказались обезьяны из рода шимпанзе (Pan). Но, как известно, род шимпанзе содержит два разных вида этих обезьян: обыкновенный шимпанзе (Pan troglodytes) и карликовый шимпанзе (Pan paniscus).
И вот интересно, что было бы, если бы в результате генетических исследований вдруг выяснилось, что озвученные виды шимпанзе произошли совсем не от какого-то общего предка, а один вид шимпанзе (Pan troglodytes) произошел, например, от гориллы (Gorilla), в то время как другой вид шимпанзе (Pan paniscus) – произошел, допустим, от человека (Homo)? При этом явное морфологическое сходство между двумя видами шимпанзе было бы предложено считать результатом конвергенции, то есть результатом независимой эволюции в сходных условиях.
Давайте посмотрим на эту картину (Рис. 21):

Рисунок 21. Слева-направо: горилла, обыкновенный шимпанзе, карликовый шимпанзе (бонобо), человек.
Интересно, что бы сказали о подобных «генетических доказательствах родства» сами свидетели Дарвина? И куда бы они послали подобные «результаты генетических исследований»?
Кстати, конфликтующие факты в вопросе о «ближайших эволюционных родственниках человека», действительно, имеются. Молекулярно-генетические исследования четко указывают на шимпанзе в качестве «ближайшего родственника». То есть, генетически, шимпанзе ближе к человеку, чем например, к горилле. В то время как по комплексу морфологических признаков человек оказывается наиболее удаленным от всех человекообразных обезьян в целом: и от шимпанзе, и от гориллы, и от орангутана (Gura, 2000).[54]
Еще один аналогичный пример конфликта генетических и морфологических фактов на уровне разных родов и видов. Как известно, сегодня к роду Пантера (Panthera) специалисты относят следующих крупнейших представителей семейства кошачьих: лев, тигр, леопард и ягуар. Эти виды крупных кошек морфологически довольно близки друг к другу. Интересно, что черепа льва и тигра (крупнейших представителей рода Pant hera) вообще трудно отличимы друг на друга.
Помимо льва, леопарда и тигра, сегодня в Старом Свете обитает еще одна крупная кошка – снежный барс (Uncia uncia). Видимо, из-за его крупных размеров, первоначально зоологи отнесли снежного барса тоже к роду пантер. Но потом разобрались, и выделили эту кошку в отдельный род (Uncia). Потому что, несмотря на сходство в размерах, морфологически и экологически (а также поведением), снежный барс серьезно отличается от всех современных представителей рода Panthera. Например, по строению черепа снежный барс отличается от всех видов рода Panthera больше, чем все они между собой в любой комбинации (Гептнер, Слудский, 1972). Более того, целый ряд морфологических и поведенческих признаков сближает снежного барса с мелкими кошками рода Felis (Гептнер, Слудский, 1972). То есть, снежный барс (род Uncia) является как бы «переходным» таксоном между крупными и мелкими кошками.
Казалось бы, если снежный барс заметно отличается от представителей рода пантер морфологически, экологически и этологически, то это означает, что он прошел больший путь независимой эволюции (по сравнению с эволюцией разных видов рода пантер от их общего предка). Следовательно, генетически, снежный барс тоже должен отличаться от пантер несколько больше, чем они друг от друга.
Однако филогенетические исследования опять преподнесли сюрприз. После их проведения оказалось, что генетически, снежный барс настолько близок к разным представителям рода Panthera, что его следует располагать прямо в их рядах. То есть, генетически, снежный барс является пантерой.
В связи с таким сюрпризом, начались серьезные попытки «впихнуть» снежного барса обратно в род пантер. Но поскольку снежный барс заметно отличается от любого другого представителя этого рода целым рядом признаков, часть специалистов пока упорно сопротивляются подобным «попыткам изнасилования» системы классификации кошек.[55]
И такие вещи (еще раз) творятся на уровне разных родов и видов, где казалось бы, всё должно быть наиболее ясно.
Последний аналогичный пример на эту же тему приведу из работы (Verheyen et al., 2003). В этом исследовании авторы на основании молекулярно-генетического анализа пытались разобраться в путях эволюции сотен видов цихлидовых рыб из африканского озера Виктория и других окрестных озер. И в результате, авторы сделали вывод, что степень генетических различий между разными видами цихлид не отражает степень их морфологических различий (Verheyen et al., 2003). То есть, генетически удаленные виды могут быть близки по внешнему виду, а генетически близкие виды могут иметь существенные морфологические отличия друг от друга.
Что уж тогда говорить об уровне семейств и отрядов. Здесь уже можно писать целые обзоры на тему «неожиданных открытий чудных». В этом удивительном мире, созданном молекулярно-генетическими исследованиями, соколиные и ястребиные птицы перестают быть близкими родственниками (Hackett et al., 2008). В то время как, например, совершенно разные (во всех отношениях) совы и птицы-мыши оказываются примерно на таком же генетическом расстоянии (друг от друга), как представители разных семейств отряда куриные (Hackett et al., 2008). Примерно такие же чудеса творятся и в мире рыб - тунцы (подотряд скумбриевидные, Scombroidei) и морские коньки (подотряд игловидные, Syngnathoidei) вдруг становятся генетическими родственниками (Betancur-R. et al., 2013). В то время как тунцы и рыбы-парусники (тоже скумбриевидные), наоборот, перестают быть родственниками, превращаясь в очередную «жертву конвергенции» (Betancur-R. et al., 2013). При этом рыбы-парусники (самые быстрые рыбы в природе), с точки зрения генетической близости вдруг уютно располагаются рядом с разными группами камбалообразных (Betancur-R et al., 2013).
У млекопитающих – тоже праздник какой-то. Генетическими родственниками становятся: сирены, слоны, златокроты, прыгунчики, трубкозубы, даманы и тенреки. Причем в наиболее близкие родственники к «водоплавающим» сиренам набиваются даманы… А сразу же за даманами идут слоны (Bininda-Emonds et al., 2007). В то время как златокроты и прыгунчики оказываются очень далеки от других насекомоядных (на которых они похожи куда больше, чем на слонов или сиренов). Бегемоты становятся генетическими родственниками китов (de Jong, 1998).
У беспозвоночных – тоже полный генетический беспредел. Многоножки вдруг становятся родственниками хелицеровым, одновременно «открещиваясь» от какого-либо родства с насекомыми… с которыми их, на самом деле, роднит целый комплекс общих морфологических признаков, из-за чего многоножек и насекомых всегда объединяли в одну общую группу (неполноусых). При этом сами насекомые оказываются настолько генетически близки к ракообразным, что некоторые особо продвинутые генетики теперь предлагают считать насекомых просто одной из групп этих самых ракообразных. Невзирая на огромные различия в морфологии и анатомии.
Впрочем, «эволюционные» деревья, построенные по генетическим данным, хотя и выглядят (нередко) забавными, но раскритиковать их на этом основании вряд ли получится. Прежде всего, потому что предполагаемая эволюция биологических таксонов – это «дела давно минувших дней» (С). Поэтому точно сказать, кто там от кого произошел на самом деле (и произошел ли) уже не представляется возможным. Даже самые неожиданные сценарии эволюционного родства, тем не менее, не исключены. Например, даже если считать стремительных тунцов родственниками неторопливых морских коньков, а еще более стремительных парусников – родственниками камбалообразных… то ведь теоретически, действительно, не исключена возможность, что за 50 млн. лет какие-то отдельные линии отклонились от своих неторопливых предков и стали эволюционировать в сторону быстро плавающих пелагических рыб. И в конечном итоге, одна такая линия породила, допустим, тунцов. То же самое независимо могло произойти еще и с предками рыб-парусников (близких к камбалообразным). И таким образом, рыбы-парусники и тунцы в итоге приобрели конвергентное сходство (хотя произошли от разных эволюционных ветвей).
Кроме того, в построенных «эволюционных деревьях», на самом деле, речь идет о степени вероятности того или иного эволюционного сценария. То есть, на конкретном «эволюционном древе» (нарисованном в очередной молекулярно-генетической работе), просто показан наиболее вероятный сценарий эволюционной истории тех или иных биологических групп, вычисленный на основе усредненного подобия исследованных генов. Поэтому если появляется какой-то факт, который совсем уж не хочет вписываться в этот эволюционный сценарий, то ничего не мешает немного изменить этот сценарий, просто перерисовав предполагаемые «ветки» и «развилки» эволюционной схемы таким образом, чтобы туда стал вписываться и новый факт тоже.
Допустим, лично мне не нравится вывод, сделанный в работе (Hackett et al., 2008), что соколиные и ястребиные птицы приобрели хищнический образ жизни независимо друг от друга (согласно молекулярно-генетическим данным). И допустим мне есть, что сказать по этому поводу. И пусть я буду даже прав. Но разве это как-то скомпрометирует «генетические доказательства эволюции»? Если в рамках этой же самой работы, сценарий единого (общего) происхождения соколиных и ястребиных птиц – тоже не исключен, а просто менее вероятен.
В общем, факты резкого расхождения между степенью генетического и морфологического (и любого другого) сходства – могут сильно удивлять, однако не опровергают молекулярно-генетические исследования как таковые.
Но можно ли тогда назвать подобные построения – «доказательствами эволюции»? Если используемая методика настолько всеобъясняющая, что способна «проглотить» вообще любые сценарии (вплоть до противоположных) - тогда какое же это «доказательство эволюции»? Это уже из серии тех «доказательств», в которых вообще всё на свете «доказывает эволюцию». На самом деле, здесь речь идет уже не о «генетических доказательствах эволюции», а о том, что в рамки эволюционной теории можно уложить почти любые генетические факты. Но между фразой «эти факты можно уложить в рамки данной концепции» и фразой «эти факты доказывают данную концепцию» - огромная разница.
Действительно, если «генетически родственные таксоны» не имеют больше ничего общего (ничего, кроме сходных генов), неужели этот факт будет «доказывать эволюцию»? В качестве «доказательства эволюции» подобные факты выглядят весьма забавно. Разве можно говорить, что молекулярно-генетические факты доказывают эволюцию, если эти факты нередко не вписываются в ранее принятые эволюционные схемы, а предлагают систематикам совершенно новые, неожиданные эволюционные сценарии, к которым этим систематикам (специалистам по соответствующим таксонам) теперь приходится привыкать. Причем некоторые предложенные генетические сценарии «эволюционного родства» оказываются настолько необычными (настолько противоречат всем другим фактам), что специалисты по конкретным биологическим группам просто не принимают от генетиков подобные схемы (несмотря ни на какое «родство генов»). И продолжают придерживаться либо прежних методов классификации, либо предлагают что-то новое, альтернативное молекулярно-генетическому подходу.
Просто в качестве конкретного примера (Расницын, 2006):
«…предлагаемый здесь тест или его более совершенные аналоги могут помочь в разрешении нынешней скандальной ситуации, когда молекулярные методы указывают на единство групп, которые по другим признакам не имеют ничего общего или почти ничего общего (знаменитые афротерии, объединение многоножек и хелицеровых и т.п.)»
Наконец, в продолжение разговора о чрезмерной пластичности «генетических доказательств эволюции», следует отметить, что примирять несоответствия между генетическими и морфологическими фактами могут еще и теоретические рассуждения о возможности разных скоростей эволюции в разных биологических таксонах.
Например, можно предположить, что один биологический таксон претерпел сравнительно быстрые генетические изменения в связи с освоением новой экологической ниши (переходом к принципиально новому образу жизни). А эволюция другого биологического таксона, наоборот, была медленной, потому что этот таксон всё время продолжал оставаться в уже освоенной им экологической нише. То есть, можно рассудить следующим образом - при быстрой смене образа жизни, возможно, те гены, которые окажутся задействованными в новом «экологическом имидже» данного таксона, будут подвергаться повышенному действию естественного отбора (и как следствие, ускоренному изменению). В то время как другие гены, не затронутые изменением образа жизни – могут и не претерпеть такого же объема изменений за то же время. В результате, данный биологический таксон, серьезно изменившись морфологически, физиологически и экологически, тем не менее, в среднем (по всей совокупности анализируемых генов), может остаться всё еще близким к какому-нибудь медленно эволюционировавшему соседнему таксону. И в итоге, допустим, быстро эволюционировавшая линия рыб-парусников оказывается всё еще генетически близкой к (теперь уже) морфологически далекой линии камбалообразных рыб (см. выше).
Однако здесь следует четко понимать, что разница в скоростях эволюции между разными биологическими группами - не может быть слишком значительной (в свете представлений о естественной эволюции, конечно). Потому что в рамках естественной эволюции, любые изменения в генах появляются случайным образом. Вследствие этого, полезные случайные мутации, необходимые для изменения соответствующих генов в нужную сторону, не могут появляться существенно чаще, чем нейтральные мутации во всех остальных генах. Полезные мутации (случайно появившись) могут только закрепляться быстрее, будучи поддержаны соответствующим естественным отбором. Но каким бы мощным ни было давление естественного отбора, всё равно, полезная мутация в определенном гене – это само по себе, весьма редкое событие. Которое происходит гораздо реже, чем нейтральные мутации, под непрерывным «дождем» из которых находятся в это время все остальные гены. Таким образом (еще раз), в рамках представлений о естественной эволюции, скорость молекулярной эволюции каких-нибудь одних генов не может намного превышать скорость молекулярной эволюции всех остальных генов (в этой же линии организмов). Так же как не могут слишком уж сильно различаться между собой и скорости эволюции в разных биологических группах.
Подобные факты гораздо лучше объясняются в рамках концепции разумного дизайна. Действительно, если произвести направленные изменения именно в тех генах, которые ответственны за новую внешность или новую биологию какой-нибудь рыбы, то и получится, соответственно, рыба с новой внешностью и новой биологией. Но в среднем (по всей совокупности генов) эта рыба всё еще будет оставаться генетически близкой к тому таксону, из которого её создали.
Так что тем верующим дарвинистам, которые пытаются объяснять, почему же генетическая близость организмов нередко конфликтует с резкими морфологическими отличиями между ними – следует быть осторожными и не постулировать слишком уж большой разницы «в скоростях эволюции» как между разными биологическими таксонами, так и между разными группами генов.
Впрочем, кажется, до сих пор никто не указал, какую же точно степень конфликта между генетическими и морфологическими фактами еще можно «простить» теории естественной эволюции, а какую – уже нельзя. И этим дарвинисты, конечно, пользуются, и продолжают уверенно списывать любые подобные факты на «неисповедимость путей эволюции».
Интересно, что уловка с постулированием «разных скоростей эволюции в разных таксонах» помогает выйти из положения не только при несоответствии генетических фактов морфологическим, но еще и в случае конфликта генетики и палеонтологии.
Вот, например, какая прекрасная фраза была написана в одной научно-популярной заметке, посвященной проблеме создания эволюционного древа млекопитающих (Наймарк, 2011):
…Как показали расчеты, скорости молекулярной эволюции в разных локусах и разных ветвях в пределах класса млекопитающих различаются не меньше чем на порядок. Именно эти специфичные для каждой ветви оценки и использовали, чтобы расположить узлы на временной оси. В результате авторы работы чуть ли не впервые получили филогенетическое древо, построенное на основе молекулярных данных, не противоречащее палеонтологическим датировкам.
Другими словами, только после тщательной подгонки теоретических «скоростей эволюции» под нужный результат, удалось (наконец-то!) согласовать молекулярно-генетические факты (степень генетических различий между разными группами) с известными палеонтологическими находками.
Если бы я захотел рассказать о том, с помощью каких хитрых приемов можно подгонять молекулярно-генетические исследования под нужные результаты – я бы, наверное, не смог выразиться яснее, чем автор только что приведенной цитаты.
Ну что же, такие приемы, действительно, способны уложить (почти любые) генетические факты в рамки почти любых палеонтологических находок. Однако после подобных откровений будет ли кто-нибудь продолжать настаивать на том, что молекулярно-генетические факты именно доказывают эволюцию?
Кроме того, я уже говорил, что при постулировании разных скоростей эволюции в разных биологических таксонах (или между разными генами) – следует иметь совесть. Потому что слишком сильный упор на такие вещи подрывает теорию естественной эволюции в целом. В рамках концепции «молекулярных часов», скорость накопления нейтральных мутаций должна быть примерно постоянной в генах разных организмов. Особенно у близких видов. И поскольку предполагается, что значительная часть молекулярных различий между генами-аналогами, на самом деле, нейтральна, то, следовательно, скорость изменения разных генов у разных видов (под действием постоянного «дождя» из разных мутаций) тоже должна быть примерно постоянной.
Тем не менее, заявления о большой разнице в скоростях генетической эволюции даже между близкими биологическими видами (!) – регулярно озвучиваются в научной печати. Точно так же, делаются заявления о разнице (иногда десятикратной!) в темпах накопления молекулярных различий между разными генами одного и того же биологического вида.
Например, у фруктовых мух из группы Drosophila melanogaster – «скорость молекулярной эволюции» гена S od оказалась в два раза выше, чем скорость эволюции этого же гена в группе Drosophila obscura. А в группе Drosophila willistoni скорость молекулярной эволюции этого же гена оказалась уже в десять раз выше (!), чем в группе Drosophila obscura (Rodriguez-Trelles et al., 2001). В то же самое время, скорость молекулярной эволюции другого гена (Xdh) в этих же группах фруктовых мух оказалась примерно одинаковой (Rodriguez-Trelles et al., 2001).
То есть (внимание) здесь мы наблюдаем одновременно и разницу в «скорости молекулярной эволюции» (десятикратную!) даже между близкими биологическими видами (принадлежащими к одному и тому же роду Drosophila), и столь же впечатляющую разницу между скоростями эволюционных изменений у разных генов в пределах генотипа одного и того же биологического вида.
Следует, конечно, понимать, что на самом деле, установленным фактом здесь является только то, что разные варианты гена S od в разных группах фруктовых мух рода Drosophila - отличаются друг от друга гораздо сильнее, чем разные варианты гена Xdh между собой у этих же видов мух. Вот это, собственно, сам факт в чистом виде. А вот заключение о разных скоростях молекулярной эволюции этих генов – это уже теоретическая интерпретация этого факта. Сделанная в рамках эволюционной концепции. Несмотря на то, что такой вывод, по сути, противоречит самой же концепции молекулярной эволюции, согласно которой (теоретически), скорости молекулярной эволюции разных генов и разных биологических таксонов должны быть примерно сопоставимы.
Напротив, в рамках концепции разумного дизайна, нет ничего удивительного в том, что одни гены-аналоги у разных организмов отличаются друг от друга в большей степени, чем другие гены-аналоги. Понятно, что при разумном конструировании генотипов разных организмов, подобная разница может быть вообще любой. Если для создания конкретного биологического вида необходимо, чтобы какие-то отдельные гены были более своеобразны, чем другие, то наверное, такое решение и следует ожидать в рамках этой концепции.
Тем не менее, авторы современных статей всё равно предпочитают объяснять подобные факты не следствием разумного конструирования соответствующих геномов, а «разницей в скоростях молекулярной эволюции». Хотя не очень понятно, какая же естественная сила могла бы порождать обсуждаемую разницу (особенно десятикратную). Постулирование «разных скоростей эволюции для разных генов» можно рассматривать, как попытку подгонки под уже принятую теорию. Где факты, не подтверждающие естественно-эволюционный сценарий, тем не менее, всё равно укладываются в рамки именно этого сценария с помощью произвольных допущений. Причем таких допущений, которые в свою очередь (сами) не очень вписываются в концепцию молекулярной эволюции.
Тем не менее, я уже говорил, что до сих пор никто не указал точно, какую же «разницу в скоростях эволюции между разными генами» – еще можно уложить в рамки естественной эволюции, а какое «изменение скорости молекулярной эволюции» - уже никак не лезет в рамки этой концепции. Поэтому такие факты продолжают интерпретироваться в русле эволюционной парадигмы и никак иначе.
Однако дела становятся совсем плохи (для дарвинистов), когда одни генетические факты начинают конфликтовать (с другими генетическими фактами) еще одним способом. А именно, когда сравнение живых существ по одним группам генов предлагает одних «генетических родственников», а сравнение этих же самых живых существ по другим группам генов – выдает уже других «генетических родственников».
Вот это уже тяжелый случай. Потому что в рамках теории эволюции, гены одного и того же организма не могут эволюционировать так, будто эти гены находились в разных биологических видах. То есть, не может организм по одним группам генов являться, например, жвачным парнокопытным млекопитающим, а по другим группам генов – змеёй (представителем чешуйчатых безногих рептилий). Когда коровы начинают «генетически дружить» со змеями (по определенным генам) – то это уже вряд ли можно назвать «генетическим доказательством эволюции». Скорее, такие факты следует назвать «доказательством разумного дизайна живых существ методами генной инженерии».
Тем не менее, согласно результатам работы (Walsh et al., 2012), коровы подружились (по определенным генам) именно со змеями.[56]
Для объяснения этого любопытного факта, авторам работы (Walsh et al., 2012) пришлось предложить тот самый «горизонтальный перенос генов», над которым мы уже посмеивались чуть выше. Причем такие горизонтальные переносы (исследованных) генов, по мнению авторов, должны были произойти не однажды, а целых девять раз! Только тогда факт удивительной близости исследованных генов у самых разных групп животных (в том числе, у коров и змей) получается «уложить» в рамки эволюционного учения (по мнению авторов данной работы).
Что и говорить, интереснейшие «доказательства эволюции» регулярно подсовывают нам молекулярно-генетические исследования. От таких «генетических доказательств» нашу бедную теорию эволюции иногда приходится просто спасать. Спасать с помощью массированного применения тех или иных дополнительных объяснений. Например, уже озвученными «горизонтальными переносами генов». Причем подобные генетические курьезы отнюдь не являются каким-то редким исключением из правил. По мере прочтения все большего числа геномов, такие факты обнаруживаются уже прямо-таки в «товарных количествах».
Вообще, это просто поразительно. Многочисленные верующие дарвинисты на всех углах продолжают уверять окружающих в том, что «генетика доказала эволюцию»… в то время как накопилось столько разных фактов филогенетических конфликтов, что уже, наверное, трудно определиться – а чего же там, собственно, больше – генетических фактов, которые ни с чем не конфликтуют (и как бы «подтверждают эволюцию»), или же генетических фактов, вступающих в разнообразные конфликты друг с другом.
То есть, на самом деле, молекулярно-генетические «доказательства эволюции» сегодня выглядят доказательствами только в сильно «причёсанном» виде – в соответствующих научно-популярных текстах и в учебниках биологии. А вот когда начинаешь читать оригинальные публикации - с завидной регулярностью обнаруживаешь там «филогенетические конфликты», в которых разные группы фактов никак не хотят «дружить» друг с другом.
Итак, давайте еще раз отметим странное поведение активных проповедников дарвинизма. Свидетели Дарвина на всех углах кричат о «генетических доказательствах эволюции», в то время как в последних научных публикациях тон несколько другой (Davalos et al., 2012):
«Несогласие среди филогений, полученных по разным наборам признаков, является распространяющимся (Rokas и др., 2003). Филогенетический конфликт становится всё более острой проблемой в связи с появлением масштабных геномных наборов данных. Эти большие наборы данных подтвердили, что филогенетический конфликт – обычен [распространен] и часто является скорее нормой, чем исключением (Waddell и др., 1999; Leebens- Mack и др., 2005; Jeffroy и др., 2006; Rodríguez-Ezpeleta и др., 2007)»
Получается, что дарвинисты-проповедники рассказывают нам о «генетических доказательствах эволюции» в то время, когда специалисты (профессионально занимающиеся этими вопросами) уже открыто признаются, что молекулярно-генетические факты чаще создают путаницу в эволюционных построениях, чем помогают чего-нибудь распутать.
И вот на этой печальной ноте давайте теперь познакомимся с таким примером «филогенетического конфликта», который резко усугубляет критичность создавшейся ситуации. Потому что, с одной стороны, этот пример тоже не вписывается в сценарий естественной эволюции, а с другой стороны, в отношении этого факта не может быть задействована даже такая спасительная «палочка-выручалочка» современного дарвинизма, как постулирование «горизонтального переноса генов». Да, именно так – сравнительно недавно «генетические доказательства эволюции» получили прямо-таки нокаутирующий удар.
Неожиданно выяснилось, что целый ряд генов у летучих мышей и дельфинов демонстрирует явную гомологию. И генов не каких-нибудь, а очень интересных. Как известно, значительная часть летучих мышей и дельфины – обладают способностью к эхолокации. И вот, чёткое сходство нуклеотидных последовательностей было выявлено сразу в 200 локусах, в многочисленных генах, связанных со слухом или глухотой. Что совместимо с предположением о причастности этих генов к эхолокации (Parker et al., 2013). Кроме того, четкие сигналы сходства были обнаружены еще и в генах, связанных со зрением.
Это уже не лезет ни в какие ворота дарвинизма. Потому что согласно современной теории эволюции, киты и летучие мыши весьма далеки друг от друга, имея родство лишь на уровне весьма древних общих предков. Отсюда следует, что эхолокация у этих таксонов должна была развиться и эволюционировать совершенно независимо. Тем более что среди родственных групп самих летучих мышей имеются виды, как способные к эхолокации, так и неспособные к ней.
Тем не менее, гены, ответственные за эхолокацию, у дельфинов и летучих мышей имеют явное сходство по большому числу участков. Настолько, что эти гены более схожи у дельфинов и тех летучих мышей, которые имеют эхолокацию, чем у летучих мышей, имеющих эхолокацию и летучих мышей, не имеющих её. Поэтому если бы какой-нибудь генетик взял бы для измерения степени родства участки ДНК, содержащие именно эти гены, то у этого генетика получилось бы, что некоторые виды летучих мышей и дельфины происходят от общего предка, в то время как другие виды летучих мышей имеют вообще других предков.
Понятно, что этот факт ну никак не ожидался в рамках дарвинизма, и прямо опровергает тезис о случайности эволюции (случайности мутаций). Потому что в рамках современной теории эволюции, крайне мала вероятность, чтобы при независимом построении сложного признака, эволюция в точности повторила одни и те же случайные мутации. Тем более, не в каком-то одном месте, а сразу во многих участках разных генов. Ну а когда речь начинает идти уже о двухстах генах (!), показывающих четкое сходство нуклеотидных последовательностей (с аналогичными генами совершенно других живых существ), то можно считать, что вывод о «неслучайности случайностей» в данном случае доказан.
Вся ирония ситуации состоит в том, что такие участки генетического сходства всегда трактовались дарвинистами, как происхождение от общего предка. Но в данном случае допускать наличие общего предка (между выборочными видами летучих мышей и дельфинами) ни в коем случае было нельзя. Потому что это значило бы вступить в очередной филогенетический конфликт. Во-первых, против целого ряда других генетических фактов, полученных на другом наборе генов, и «говорящих нам», что дельфины и летучие мыши – не имели близких общих предков. Во-вторых, еще и против явного морфологического различия между летучими мышами и дельфинами. И в-третьих, против общепринятых схем эволюции этих биологических таксонов.
Пресловутым «горизонтальным переносом» этот факт тоже не объяснить. Потому что обнаруженное генетическое сходство имеет чересчур массированный характер. Здесь сходство демонстрируют сразу сотни генов, причем не абы какие, а именно те, которые ответственны за обеспечение слуха. Поэтому если бы в данном случае кто-нибудь решил выдвинуть версию о случайном «горизонтальном переносе», то пришлось бы объявлять миру об открытии горизонтального переноса уже не отдельных генов, а о переносе сразу всех генов слухового аппарата. Понятно, что такой случайный «горизонтальный перенос» (слуховых аппаратов сразу целиком) от одного животного к другому – это фантастика чистой воды. Поэтому версию «горизонтального переноса генов» в данном случае никто и не озвучил.
Какой же выход нашли из всего этого дарвинисты? Может быть, они отказались от своих теоретических эволюционных схем, придя к выводу, что с этими схемами что-то не так? Нет, что Вы. Разве такое вообще возможно?
Верующие дарвинисты просто сделали следующее заключение:
- Ах, оказывается, одни и те же вызовы среды всё-таки могут закреплять одни и те же мутации в одних и тех же генах! (даже если этих генов – две сотни).
То есть, естественный отбор, оказывается, обладает способностью «отлавливать» именно одинаковые мутации, чтобы в совершенно независимых эволюционных случаях создавать сложные признаки одинаковым генетическим способом. Даже столь серьезный признак, как способность к эхолокации, «завязанный» сразу на множество генов. Видимо потому, что «гипотетическое пространство возможных решений» для возникновения соответствующего признака настолько строго ограничено, что здесь могли закрепляться только строго определенные случайные мутации (а другие вообще не закреплялись). Еще раз отметим, что подобные вещи до этого считались невероятными (и правильно считались), а вот теперь сразу стали считаться вероятными.
На самом деле, факт столь удивительной «генетической конвергенции» объясняется в миллион раз правдоподобнее, если признать, что за формирование сложных биологических устройств ответственны отнюдь не случайные мутации, а что-нибудь более предсказуемое. Например, разумный дизайн. Где сходство генов, ответственных за эхолокацию (у столь разных организмов, как летучие мыши и дельфины), объясняется просто применением сходных инженерных решений при создании сходных функциональных устройств.
Поразительно, но дарвинисты эту гипотезу (столь очевидно напрашивающуюся в данном случае) почему-то вообще отбрасывают, да еще и обзывают псевдонаукой. И предпочитают считать подобные совпадения (тысяч нуклеотидов) результатом работы не разумного «генного инженера», а следствием одинакового естественного отбора, произведенного на одни и те же гены, в которых произошли одни и те же случайные мутации. При этом они продолжают спокойно называть себя учеными, а не разносчиками невероятных небылиц.
Но самое смешное здесь в другом. Дело в том, что биологи, обнаружившие факт совпадения (специфических) генов у совершенно разных организмов, и решившие объяснить этот большой сюрприз результатом «обширной генетической конвергенции»… тем самым нанесли страшный удар по своим «братьям меньшим» – по дарвинистам-проповедникам, активно проталкивающим свой дарвинизм в широкие народные массы. Разнообразные «народные просветители», кажется, уже уверили народ в том, что генетические факты всё-таки доказывают происхождение биологических таксонов от общих предков… И тут авторы статьи о летучих мышах и дельфинах, во-первых, «обрадовали» самим фактом установления «генетической конвергенции», а во-вторых, предположили, что этот феномен, может быть, гораздо шире распространен в природе, чем принято было думать (Parker et al., 2013).
Ну и что теперь прикажете делать среднему дарвинисту-проповеднику? Ведь теперь практически любой факт генетического сходства – уже не является серьезным доказательством именно «эволюции от общего предка». Ибо этот факт может оказаться отнюдь не «свидетельством происхождения от общего предка», а просто следствием «одинакового давления естественного отбора, произведенного на одни и те же гены в строго ограниченном пространстве возможных мутаций». То есть, следствием генетической конвергенции (или генетического параллелизма). В общем, попробуй теперь разберись, где здесь генетически доказанная «эволюция от общего предка», а где просто результат «генетической конвергенции».
Более того, после такого страшного удара по молекулярной филогении, к дарвинисту-проповеднику, рассказывающему о том, что «генетика уже доказала эволюцию», теперь ведь обязательно подойдет какой-нибудь коварный креационист, и нагло заявит, что никаких «общих предков, доказанных генетическими исследованиями», на самом деле, не существовало вовсе. А есть лишь факт наличия «генетической конвергенции». Который можно, конечно, объяснить тем, что «одинаковое давление естественного отбора отбирало одни и те же случайные мутации в одних тех же генах»... но гораздо правдоподобней объяснить этот же самый факт - результатом применения сходных инженерных решений при создании сходных функциональных устройств (каким-то разумным агентом).
А теперь давайте вспомним еще и про другие филогенетические конфликты, на которые уже натолкнулась филогенетика. Причем (еще раз) филогенетика стала наталкиваться на такие факты всё чаще. Тем чаще, чем больше генетических данных становится доступно для анализа.
Например, при исследовании генома бделлоидной коловратки[57] вдруг оказалось, что помимо генов животных, у неё присутствуют: 1) гены бактерий, 2) гены растений, 3) гены грибов (Gladyshev et al., 2008). Понятно, что проще всего это интерпретировать, как явное свидетельство прямого дизайна организмов (методами генной инженерии?). Однако дарвинисты предпочли списать такие вещи на «массовый горизонтальный перенос генов». То есть, коловратки просто взяли и заимствовали (самостоятельно) сотни генов у растений, грибов и бактерий, причем заимствовали способом, пока неизвестным современной науке. Видимо, коловратки - это такие талантливые генные инженеры. Интересно, что авторы оригинального исследования, похоже, сами испугались своих удивительных результатов, и бросили это дело, проанализировав всего 1% генома бделлоидной коловратки. А жаль. Сколько еще «открытий чудных» (наверное) готовили явить миру остальные 99% генома коловратки.
И вот теперь ко всему этому филогенетическому «беспределу» добавилась еще и «генетическая конвергенция».
Хотя на самом деле, «генетическая конвергенция» начала поражать биологов несколько раньше. Например, ранее уже была установлена удивительная «генетическая конвергенция» между механизмом, обеспечивающим формирование речи у человека, и механизмом, обеспечивающим способность к пению у певчих птиц. Оказалось, что и в том и в другом случае развитие этих способностей опирается на одно и то же молекулярно-генетическое основание, связанное с геном FOXP2 (Bolhuis & Okanoya, 2010). Несмотря на то, что в рамках дарвинизма, эволюция этих механизмов у людей и птиц должна была протекать совершенно независимо. Между тем, генетические механизмы «по странной случайности» оказались сходны. Причем сходство здесь имеется не только на генетическом уровне, но еще и на анатомическом – наблюдается удивительное сходство в анатомии соответствующих зон мозга (Bolhuis & Okanoya, 2010). Кроме того, освоение речи человеческими младенцами и освоение пения птенцами певчих птиц выглядит сходным еще и на поведенческом уровне (Bolhuis & Okanoya, 2010).
Наконец, в совсем уже недавнем исследовании выяснилось, что ген этого же семейства (dFoxP) каким-то образом задействован еще и в высшей нервной деятельности плодовых мушек дрозофил (DasGupta et al., 2014). Причем, похоже, задействован этот ген у плодовых мушек именно там, где мухе необходимо «общаться» с окружающим миром и принимать решения.
Но ведь членистоногие – настолько далеки от позвоночных, насколько вообще может быть удалено одно билатеральное животное от другого. Получается, что либо «последний общий предок» всех билатеральных животных уже был весьма сообразительным парнем, либо это очередное «случайное эволюционное совпадение». Либо (самая простая гипотеза) это результат сходного дизайна разных биологических таксонов.
Кстати, так получается не только по этим генам, но еще и по многим другим. Например, совсем недавно выяснилось, что химические механизмы, ответственные за возникновение тревоги в нервной системе раков и позвоночных животных - тоже одинаковы (Fossat et al., 2014).
Нашлись «случайные эволюционные совпадения» и между нервной системой круглых червей и позвоночных. Конкретно, химический механизм, отвечающий за процессы обучения и полового поведения круглого червя Caenorhabditis elegans, оказался сходен с аналогичным механизмом, играющим важную роль в регуляции поведения (особенно полового и социального) у млекопитающих (Марков, 2012а).
Столь же интригующую информацию выдала нам еще одна научно-популярная заметка (Марков, 2012б). Оказывается, гены, склоняющие психику (того или иного животного) к поиску чего-нибудь новенького… являются сходными у пчел и млекопитающих. Условно назовём эти гены – «генами любопытства». Так вот:
…Самое интересное, что эти гены связаны с поиском новизны также и у позвоночных. Так, в мозге пчел-разведчиц понижена экспрессия гена DopR1, кодирующего дофаминовый рецептор первого типа (D1). У крыс с повышенной тягой к новизне тоже отмечена низкая экспрессия гена рецептора D1 (Viggiano et al., 2002)…
Действительно, очень интересно. Опять получается одно из трех:
1. Или (в свете «единственно верного учения») общий предок всех билатеральных животных был уже не только сообразительным, но и весьма любознательным парнем. И еще он мог тревожиться (по разным поводам). А также обучаться.
2. Или это всё снова «чисто случайные эволюционные совпадения».
3. Или же это хорошие свидетельства сходного дизайна (нервной системы беспозвоночных и позвоночных животных).
Заключение, сделанное автором заметки (глубоко верующим дарвинистом Александром Марковым), следует, наверное, привести прямо отдельными фразами (с комментариями):
«Тот факт, что нейрохимические основы поискового поведения оказались сходными у насекомых и млекопитающих (включая человека), заставляет задуматься о закономерностях эволюции поведения у животных в целом…»
Да уж, заставляет.
«…Последний общий предок пчел и людей жил, по-видимому, 650–700 млн лет назад, в криогеновом периоде. Не исключено, что у этого предка, который, возможно, был похож на сегментированного червячка с парными придатками, уже было что-то похожее на поисковое поведение и дофаминэргическую систему вознаграждения»
А еще, судя по сходству других групп генов (между совершенно разными биологическими таксонами животных), этот загадочный общий предок, помимо развитой нервной системы с механизмами высшей нервной деятельности, тревоги, внутреннего вознаграждения, стремлением к новизне и способностью к обучению… имел еще и глаза, сердце, конечности, половое поведение, механизм развития через метаморфоз и т.п. (см. ниже). В связи с этим хочется спросить, а не курил ли этот «общий предок» еще и трубку?
Продолжаем цитату:
…Может быть, более сложные формы такого поведения вместе с похожими системами его регуляции развивались параллельно на общей, унаследованной от далеких предков нейрохимической основе. Любопытно, что даже у круглого червя C. elegans недавно обнаружена связь между особенностями поведения при поиске пищи и аллельными вариациями гена тираминового рецептора (Bendesky et al., 2011). Тирамин — нейромедиатор из группы катехоламинов, близкий к дофамину и октопамину…[58]
Действительно, любопытно. Значит, сходство с круглыми червями имеется еще и в этом отношении.
…Не выяснится ли в конце концов, что нейронные механизмы, ответственные за принятие решений, мотивацию поведения и даже эмоции, сходны в своей основе у всех билатерально-симметричных животных?
Но когда это выяснится, этот поразительный факт, конечно же, всё равно не будет свидетельствовать в пользу единого дизайна животных. Из этого факта всё равно сделают вывод, что «учение Дарвина всесильно, потому что оно верно».
Если же говорить серьезно, то нервная система членистоногих и позвоночных анатомически различается очень сильно. Отсюда, в рамках самого же эволюционного учения неизбежно следует вывод, что общий предок этих животных мог иметь только очень примитивную нервную систему. Допустим, как у гидры, у которой нервная система представляет собой сеть разбросанных по телу нервных клеток. Или что-нибудь немного более сложное. Например, как у круглых червей. И уже от этого простого исходного состояния, нервная система членистоногих постепенно эволюционировала в одну сторону, а нервная система позвоночных – в другую. При этом нервные системы обоих таксонов в ходе воображаемой эволюции постепенно (и независимо!) усложнялись, вплоть до появления элементов высшей нервной деятельности – специальных механизмов принятия решений и «общения» с миром, механизмов, обеспечивающих любопытство, реакции тревожности и прочее. Казалось бы, в ходе случайной эволюции, эти механизмы в конечном итоге должны были получиться биохимически разными, просто в силу фактора случайности (мутаций). Но нет, эти механизмы вдруг (!) оказались сходными. Если мы не хотим совершать революцию в математике, опровергая теорию вероятностей, но при этом желаем оставаться в рамках дарвиновского учения, нам придется заключить, что членистоногие и позвоночные просто унаследовали механизмы высшей нервной деятельности от некоего общего предка, чья нервная система уже была достаточно совершенной, обладая всеми перечисленными выше свойствами. Но в таком случае мы вступаем в противоречие с только что сделанным (нами же) заключением об исходной примитивности нервной системы этого общего предка, которое следует из наблюдаемых анатомических различий между нервными системами членистоногих и хордовых.
Таким образом, в рамках «единственно верного учения» мы получаем очередную «загадку эволюции». Что не может не забавлять, потому что в свете другой биологической концепции – теории разумного замысла – эта «великая загадка» вообще не представляет никаких проблем, а наоборот, весьма ожидаема. В рамках концепции разумного замысла, подобное сходство объясняется просто сходными творческими решениями, которые были применены при создании одних и тех же вещей (механизмов нервной деятельности) в разных биологических таксонах.
Всё только что сказанное касается и кровеносной системы. Потому что кровеносные системы позвоночных и членистоногих тоже различаются радикальным образом. Тем не менее, определенные гены, отвечающие, например, за развитие сердца – явно сходны друг с другом (Scott, 1994). Несмотря на то, что строение сердца членистоногих радикально отличается от строения сердца позвоночных. Что, казалось бы, свидетельствует о независимом эволюционном происхождении этих сердец.
И наконец, самый поразительный пример – это общность механизмов построения глаза. Дело в том, что у животных из разных биологических таксонов глаза настолько различаются по своему анатомическому строению и принципам работы, что большинство специалистов всегда искренне считало - глаза в ходе эволюции появились независимо в разных эволюционных линиях животных. Возможно, более 40 независимых раз подряд (Жуков, 2010).
Однако результаты генетических исследований поразили всех так, что шум до сих пор не утихает. Например, оказалось, что за «разметку» глаз у членистоногих, позвоночных и даже кишечнополостных (!) отвечают сходные гены из группы Pax.[59] Что представляется фактом, достойным всяческого удивления. Действительно, почему из всего имеющегося разнообразия факторов транскрипции, на роль «разметчика» глаз были выбраны именно гены группы Pax в трех независимых эволюционных случаях? Причем эти мастер-гены оказались настолько сходны, что Pax-6 мыши, «пересаженный» мухе, привел к образованию глаза у этой мухи.
Еще более удивительно, что сходные гены включают формирование глаз еще и у медуз! Конкретно, ген Pax-A отвечает за формирование глаз у гидроидных медуз (Suga et al., 2010). А ген Pax-B «запускает» глаза кубомедузам (Kozmik et al., 2003). Причем эти гены, опять-таки, оказались настолько близки к генам Pax других животных, что Pax-А гидроидной медузы (так же как и Pax- B кубомедузы), «встроенные» в муху, столь же добросовестно вырастили глаза на теле этой мухи (Suga et al., 2010; Kozmik et al., 2003), как и Pax-6 мыши в более раннем исследовании.[60]
И это вообще из ряда вон, потому что медузы – это даже не билатеральные животные (медузы являются радиально-симметричными существами). Представить себе общего предка медузы и билатеральных животных уже глазастым – весьма сложно. Ведь считается, что билатеральные животные произошли от радиально-симметричных чуть ли не на самых ранних этапах эволюции животных, когда представители животного царства были устроены еще очень просто. Тем не менее, глаз кубомедузы «взорвал» теорию эволюции не только общим геном Pax, но еще и сходными белками опсинами, которые оказались даже более близки к опсинам позвоночных, чем, например, к опсинам членистоногих. А ведь кишечнополостные на официальном «эволюционном древе» удалены от позвоночных существенно дальше, чем членистоногие (членистоногие вместе с позвоночными относятся к билатеральным животным). Тем не менее, опсины кишечнополостных, наоборот, ближе к опсинам позвоночных, чем опсины членистоногих. В рамках теории эволюции такого, вроде бы, не может быть. Не может быть потому, что глаза медуз не могли эволюционировать отдельно от самих медуз (примкнув в этом деле к позвоночным). Именно такие «чудеса в решете» сейчас и называются филогенетическими конфликтами. И для объяснения подобных чудес выдумываются специальные теоретические схемы (под каждый отдельный случай), которые позволяют всё-таки объяснить, почему так произошло в рамках дарвиновской теории. Например, конкретно по поводу конфуза с опсинами позвоночных и медуз было выдвинуто предположение, что это они просто так «хорошо сохранились», то есть, опсины позвоночных по какой-то причине остались ближе к «обще-предковым» опсинам, чем опсины членистоногих.[61]
Но у кубомедуз имеется еще одно вещество, задействованное в глазу, которое тоже оказалось сходным с соответствующим веществом позвоночных животных. Это меланин (Kozmik et al., 2008). И выполняет он в глазу кубомедузы (уже догадались?) точно такие же функции, что и в глазу позвоночных.[62] Хотя если бы речь шла о независимом развитии глаза кубомедузы в ходе случайной (!) эволюции, то на начальных этапах такой эволюции место меланина могло бы, наверное, занять практически любое непрозрачное вещество, вырабатываемое живой клеткой (например, у насекомых аналогичную функцию выполняют уже другие пигменты – оммохромы). Тем не менее, это вещество оказалось отнюдь не любым, а именно меланином. То есть именно тем, что имеется в глазу позвоночных, и выполняет там такие же функции.
Более того. Оказалось, что еще и регуляция синтеза меланина в глазу кубомедузы осуществляется тем же транскрипционным фактором (MITF), что и у позвоночных (Kozmik et al., 2008).
Таким образом, глаза кубомедуз и позвоночных оказались близки по целому ряду генетических признаков (гены группы Pax, гены меланогенеза, гены опсинов). Да и морфологически, камерные глаза кубомедуз тоже напоминают глаза позвоночных животных во многих отношениях (Жуков, 2010).[63] То есть, глаз медузы имеет целый комплекс сходных признаков с глазом позвоночных - начиная от чисто поверхностного сходства (на уровне общих аналогий) до весьма глубокого.
Поэтому ночной кошмар с глазастым общим предком всех животных уже прямо-таки нависает над нами (в рамках теории эволюции):
Рисунок 22. Вверху – одна из воображаемых схем эволюционного появления билатеральных животных из древнейших радиально-симметричных предков. Красным цветом выделен аборальный нервный центр. Взято из (Малахов, 2004). Внизу – та же самая картинка, но уточнённая соответствующим образом - все воображаемые «общие предки» должны были быть уже глазастыми.
И наоборот, этот «глазастый кошмар» сразу же отступает, как только мы попробуем рассмотреть эти же факты с позиций разумного дизайна живых существ. В этом случае, множественные параллели между глазами столь разных организмов перестают вызывать какое-либо недоумение, представляясь вполне обычным следствием сходных «технических» решений, реализованных в сходных живых «конструкциях».
Более того, в свете этой же концепции одновременно получают объяснение и резкие различия в строении глаз этих животных, которые тоже имеются, несмотря на целый ряд сходных признаков. С позиций представлений об «общем предке» - непонятно, почему строение глаз разных животных настолько (принципиально) разное. Если мы предполагаем, что свои глаза все эти животные унаследовали уже в готовом виде.[64]
А вот в свете концепции разумного дизайна, здесь тоже всё понятно. Потому что глаза разных животных вполне могли быть сконструированы очень по-разному, но с использованием некоторых сходных элементов (конструкторских решений).
Но и это еще не всё. Глаз кубомедузы неожиданно оказался весьма совершенным, но это совершенство медузе не нужно, потому что у неё просто нет возможностей, чтобы этим совершенством воспользоваться (Nilsson et al., 2005; Веденина, 2006). А именно, хрусталик кубомедузы обладает превосходными оптическими свойствами и за счет отличных преломляющих свойств передает изображение почти без искажений.[65] Таким образом, хрусталик кубомедузы способен был бы обеспечивать весьма качественное изображение… если бы только: 1) нервная система медузы могла обработать столь качественную картинку (а нервная система медузы не может этого сделать из-за своей примитивности) и 2) если бы фокусное расстояние от хрусталика до сетчатки в глазе кубомедузы не было бы сильно сбито (Nilsson et al., 2005). С таким сбитым фокусным расстоянием даже человек видел бы мир лишь в виде туманных образов.
Получается, что хрусталик медузы может обеспечить качественное и детализированное изображение. Но медуза использует его лишь для того, чтобы (по одной из гипотез)… различать столбы света в морской воде! Это всё равно, как если бы Вы, имея микроскоп, использовали его только для того, чтобы определить, темно в комнате или нет. Действительно, таким способом использовать микроскоп, в принципе, можно.
Получается, что свои (потенциально) хорошие глаза, кубомедузы используют больше в качестве украшения, чем по прямому назначению.
Ну что же, глаза у кубомедузы, действительно, довольно симпатичные. Так может быть, эти глаза - результат полового отбора? Иначе как еще можно объяснить появление под действием естественного отбора такого органа, возможности которого никак не используются самим организмом (и не могут быть использованы в принципе).
Так что глаза кубомедуз (наверное) эволюционировали в ходе придирчивого полового отбора, когда одна медуза выбирала другую медузу с самыми симпатичными глазами (и самыми качественными хрусталиками). Правда, делали они это наощупь, потому что медузы физически не способны различать такие детали.
Не менее интригующим было обнаружение множественного сходства в генетических механизмах развития конечностей позвоночных и крыльев насекомых на ранних стадиях. Здесь уже целый комплекс сходных мастер-генов занимается сходной работой в сходных местах (Шаталкин, 2003). Несмотря на то, что крылья насекомых и конечности позвоночных, по единодушному мнению специалистов, произошли независимо друг от друга.
Привожу соответствующую цитату (Шаталкин, 2003:79). Рекомендую набраться сил и всё-таки одолеть эту цитату полностью, несмотря на то, что текст густо пересыпан научными терминами:
…Наличие гомологичных (ортологичных) генов, определяющих эквивалентные морфологические структуры у далеких в филогенетическом отношении групп, вновь поставило перед морфологами проблему гомологии, казалось бы, уже окончательно решенную. Суть проблемы в том, что негомологичные органы (например, крылья у насекомых и птиц) могут определяться сходным комплексом гомологичных генов…
...С этой точки зрения особенно поражает онтогенетическое сходство крыльев [насекомых] и конечностей тетрапод на начальных фазах их формирования. В частности, показана (Wolpert et al., 1998) четкая параллель экспрессии гомологичных генов между апикальным эпидермальным гребнем (АЭГ) у цыпленка и аналогичного по функции крылового края у Drosophila. В обоих случаях эти структуры разделяют вентральный и дорсальный компартменты, причем в клетках последнего имеет место экспрессия Radical fringe (R-fng) у цыпленка или его гомолога fringe (fng) у дрозофилы. Индуктивный эффект fng в крыловом крае опосредуется через лиганды Delta и Serrate, запускающие через рецептор Notch соответствующую сигнальную систему. Аналогично этому, в крыле цыпленка индуктивный эффект R-fng в АЭГ также опосредуется через гомологичные белки Serrate-2 и Notch-1.
К этому следует добавить, что имеется интересная функциональная параллель между АЭГ у цыпленка и краевыми клетками крылового имагинального диска. Подобно АЭГ у цыпленка, крыловой край у Drosophila необходим для дистального развития крыла, и частичная его потеря в обоих случаях ведет к дефектам вдоль проксимо-дистальной оси развивающихся крыльев.
В крыловом крае у дрозофилы секретируется морфоген Wingless. Его гомолог Wnt-7 отмечен у цыпленка в дорсальной эктодерме крыловой почки (рис. 16). Еще одна параллель связана с образованием передне-задней границы в крыле. У дрозофилы в задней части будущего крыла проявляет активность ген hedgehog, и его гомолог Sonic hedgehog также активен в задней области крыловой почки цыпленка. На передне-задней границе крылового зачатка дрозофилы происходит выделение морфогена Decapentaplegic. Его гомолог BMP-2 обнаруживается в задней трети крыловой почки цыпленка, т.е. не имеет, в отличие от дрозофилы, краевого распространения. У дрозофилы с дорсальной областью будущего крыла связан ген apterus. Его гомолог в крыловой почке цыпленка Lmx-1 экспрессируется в дорсальной мезодерме.
Очевидно, что параллелизмов очень много, и их нельзя объяснить независимой кооптацией туловищных генов при формировании конечностей. Какие же еще могут быть правдоподобные объяснения столь многочисленных параллелизмов в группах, никак не связанных между собой?
Итак, мы наблюдаем множественные генетические совпадения в развитии таких органов, которые сходны по выполняемой функции, но совершенно разные по происхождению. Чтобы всё-таки уложить этот удивительный факт в рамки «единственно верного учения», специально для этого случая было выдвинуто несколько гипотез (см. Шаталкин, 2003:80).
В свете дальнейшего обсуждения «генетических доказательств эволюции», интересно также, что у насекомых генетические механизмы развития крыльев отличаются от механизмов развития их же ног (Шаталкин, 2003):
Развитие крыльев насекомых идет, как мы видели, по несколько иной схеме. Это, возможно, свидетельствует о существовании у насекомых двух различных механизмов эволюции ног и крыльев.
Получается (немного иронизируя над «генетическими доказательствами эволюции»), что ноги и крылья насекомых происходят от разных предков. Ноги - от какого-то неизвестного «общего предка всех насекомых»,[66] а вот крылья, судя по генетическим параллелям, имели общего предка с позвоночными животными?
Хотя при этом заранее известно, что никаких общих предков именно в деле формирования крыльев – у позвоночных с насекомыми быть не могло. Не могло просто потому, что первые насекомые были вообще бескрылыми (в рамках представлений самой же теории эволюции).
Кстати, разные группы первичнобескрылых насекомых живут припеваючи и по сей день. Поэтому любой интересующийся может сам оценить, какие именно «зачатки крыльев» (доставшиеся от общих предков с позвоночными?) могли иметься у первых насекомых. Для этого необходимо заглянуть в какой-нибудь укромный влажный уголок и обнаружить там вот такое создание (Рис. 23):

Рисунок 23. Обыкновенная чешуйница (Lepisma saccharina), отряд щетинохвосток (Thysanura), первичнобескрылые насекомые.[67]
Как видим, никаких следов крыльев. Это представительница щетинохвосток, которые являются первичнобескрылыми насекомыми. Помимо щетинохвосток, первичнобескрылыми являются и другие группы шестиногих (Hexapoda): бессяжковые (Protura), двухвостки (Diplura) и ногохвостки (Collembola). Однако они относятся к группе скрыточелюстных (Entognatha), то есть, не совсем к насекомым (в узком смысле), а к древней группе живых существ, которые являются наиболее близкими к насекомым и как бы намекают на то, как могли бы выглядеть эволюционные предки современных насекомых.[68] А вот щетинохвостки (Thysanura) – относятся уже к настоящим насекомым (Insecta), которых объединяет большое число общих признаков (строение челюстей, усиков и многое другое).[69]
Тем не менее, щетинохвостки являются первичнобескрылыми. В связи с этим не очень понятно (в рамках «единственно верного учения»), откуда вдруг в крыльях насекомых (тех, у которых имеются крылья) взялись множественные генетические параллели… с развитием конечностей позвоночных животных!
Но и это еще не всё. Другая (небольшая) генетическая параллель была установлена в отношении «ушей и носов» насекомых и позвоночных (Шаталкин, 2003):
…Ген Distalless является ключевым в образовании дистальных частей конечностей (Panganiban, 2000; Schoppmeier & Damen, 2001), и на нем следует остановиться подробнее. Этот ген имеет несколько функций. У дрозофилы он проявляет себя в антеннах, в которых, помимо определения дистального компартмента, играет роль в развитии слуховых и обонятельных рецепторов. Интересно, что у позвоночных ген Dlx, являющийся гомологом Dll, также имеет значение в развитии слуховых органов и носа, и именно эта функция, как предполагают, была исходной (Panganiban & Rubenstein, 2002).
Еще одна серия интересных генетических параллелей была установлена уже не между насекомыми и позвоночными, а между знаменитым утконосом и некоторыми группами других позвоночных животных. Как известно, утконос – весьма удивительное создание, одновременно сочетающее в себе черты млекопитающих, птиц и пресмыкающих. И вот, в ходе соответствующего исследования оказалось, что такие же параллели прослеживаются еще и на генетическом уровне (Warren et al., 2008).
Например, известно, что у самцов утконоса на задних ногах имеются хорошо развитые ядовитые роговые шпоры, при помощи которых можно отправить на тот свет некрупное животное вроде собаки. Эти шпоры имеют самые настоящие ядовитые протоки, которые, в свою очередь, связаны с самой настоящей ядовитой железой, находящейся у утконосов тоже в ногах. Таким образом, здесь имеется очевидная аналогия с ядовитым аппаратом змей. Только у змей весь их ядовитый аппарат приурочен к голове и клыкам, а у утконоса – к ногам. И вот, в результате проведенных исследований (Warren et al., 2008) было установлено, что химический состав яда утконоса обеспечивается генами, которые… тоже имеют сходство с соответствующими генами змей! В связи с этим поразительным открытием (очередной «генетической конвергенции») авторы работы написали, что в ходе независимого эволюционного формирования ядовитого аппарата змей и утконосов, гены, отвечающие за состав ядов, были независимо (!) избраны из тех же самых генных семейств![70]
Хотя, казалось бы, почти бесконечное разнообразие ядов, которое имеется в живой природе, красноречиво указывает на возможное число вариантов. Что и говорить, если бы я обладал удачей (в отношении чисто случайных событий) в той же степени, какую всё время демонстрирует нам «случайная эволюция аналогичных органов» - я бы, наверное, уже выиграл в лотерею миллион рублей 20 раз подряд.
И наконец, по поводу развития через метаморфоз.
Оказалось, что запуск механизма метаморфоза у очень далеких биологических групп – медуз, насекомых, позвоночных - тоже осуществляется с помощью сходной сигнальной системы (Fuchs et al., 2014). Но ведь развитие через метаморфоз – это, скорее всего, не предковое (не исходное) состояние этих биологических таксонов (максимально далеких друг от друга). Тем не менее, оказалось, что сигнальные системы, запускающие процесс метаморфоза, у всех перечисленных таксонов имеют сходный характер.
Вот что пишет по этому поводу всё тот же Александр Марков, которому как-то ведь надо «объяснять» столь удивительные «совпадения» именно в рамках «единственно верного учения» (Марков, 2014):
…Случайное совпадение, конвергенция или свидетельство общего происхождения? Для ответа на этот вопрос данных пока недостаточно.
Таким образом, механизмы запуска генетической программы метаморфоза у позвоночных, насекомых и книдарий оказались на удивление сходными. Возможно, это говорит о том, что уже у общего предка этих групп (то есть у последнего общего предка Eumetazoa) был сложный жизненный цикл с метаморфозом. Или, может быть, метаморфоз у них развился независимо, но во всех случаях для его регуляции был привлечен один и тот же ретиноевый сигнальный каскад, имевшийся у общего предка, но служивший ему для иных целей (для включения других наборов генов).
Бедный Александр Марков всё никак не может решиться, и написать предположение, которое в данном случае является самым очевидным:
…Случайное совпадение, конвергенция, свидетельство общего происхождения… или следствие сходного дизайна?
Вот что должен был бы написать Александр Марков по этому поводу.
Но не напишет он так никогда. Потому что «обратного пути уже нет» и «мосты сожжены». И если где-нибудь обнаружат медузу, у которой в генах будет расшифрована даже прямая подпись её разумного создателя, Марков отвернется от этой подписи, и начнет рассуждать, что же это было – «случайное совпадение, результат конвергенции или эволюция от общего предка»?
Но если продолжать упираться таким образом, рассматривая всё новые обнаруживающиеся параллели (между совершенно разными животными) только в рамках естественной эволюции и никак иначе, то мы получаем интересную тенденцию в наших теоретических схемах. Интересно наблюдать, как привычное нам (со школьной скамьи) «древо эволюции» под давлением научных открытий постепенно перестаёт быть «деревом». А превращается в совершенно откровенный «куст». Где чуть ли не все основные гены, органы и даже системы органов, имеющиеся у современных животных… уже имелись у некоего (весьма таинственного) последнего предка всех животных. Который (воображаемо) «сидел» в самом основании этого «эволюционного куста». А потом по-быстрому эволюционно разветвился, породив разные типы многоклеточных животных.
То есть, под давлением фактов, верующие дарвинисты вынуждены всё больше повышать теоретический возраст возникновения основных групп генов, ответственных у животных за развитие разных органов и систем органов. Помещая эти гены во всё более «древнюю древность». Уже чуть ли не в «первичный бульон».
Причем палеонтология такое «явление Христа народу»… пардон, появление общего предка всех животных, столь основательно нагруженного сразу всеми необходимыми генами и органами - отследить пока не в состоянии. Какие-либо фактические свидетельства эволюции, в ходе которой постепенно появлялись бы сотни разных генов, необходимых для построения разных органов животных… не установлены палеонтологической летописью. Вместо этого установлено внезапное «выпадение из сингулярности» практически всех известных сегодня типов животных во время так называемого «кембрийского взрыва». Причем животные разных типов «выпадают из сингулярности» сразу с готовыми глазами, мозгами и сердцами, без каких-либо палеонтологических свидетельств предшествующей эволюции этих органов (см. ниже).[71]
В общем, мы сейчас, похоже, наблюдаем постепенный крах аргумента про «генетику, уже доказавшую эволюцию» прямо в режиме реального времени. Довольно быстро накапливаются факты многочисленных «генетических конвергенций» между биологическими таксонами разного уровня. Начиная от «генетической конвергенции» между разными типами животных (последние примеры), и кончая «генетической конвергенцией» в пределах разных отрядов или даже семейств (как в озвученных выше случаях с белками-антифризами рыб или эхолокацией дельфинов и летучих мышей).
Если такие милые сюрпризы с массированной «генетической конвергенцией» продолжат стабильно обнаруживаться и дальше, то разнообразным «народным просветителям», посвятившим долгие годы проповедям дарвинизма среди населения с помощью именно «фактов генетического родства», наверное, придётся просто застрелиться от стыда.
Хотя какое-то не слишком человеколюбивое напутствие я сейчас высказал. Надеюсь, это напутствие никто всерьез не воспримет. Даже самый совестливый верующий дарвинист. Я призываю всех верующих дарвинистов – после того как окончательно выяснится, что «генетические доказательства эволюции», на самом деле, никуда не годятся – не впадайте в депрессию и не стреляйтесь. Примите этот факт с мужеством, и продолжайте жить. Ведь помимо «генетических доказательств эволюции», существуют еще и другие «доказательства». Например, Вы можете продолжать «доказывать эволюцию» с помощью ссылок на выведение разных пород собак и голубей. Чем бы дитя ни тешилось, лишь бы не плакало.
Наконец, в контексте завершения разговора о филогенетических конфликтах, интересно узнать, что обнаружены и такие факты, которые по своему смыслу прямо противоположны озвученным выше, но тем не менее, тоже являются филогенетическими конфликтами.
Выше мы увидели, что вопреки эволюционным ожиданиям, совершенно неродственные таксоны (чрезвычайно удаленные друг от друга на «древе жизни» в рамках современной теории эволюции), тем не менее, почему-то обладают сходными генами, выполняющими сходные функции. Или даже целыми комплексами сходных генов.
Однако современная генетика установила и такие факты, когда наоборот, родственные таксоны используют совершенно разные гены. Причем не где-нибудь, а на фундаментальных этапах собственного развития.
Например, в работе (Klomp et al., 2015) неожиданно обнаружилось, что за формирование передне-задней оси тела у мух дрозофил и комаров-звонцов отвечают разные гены. Причем это различие состоит отнюдь не в единственном гене, а в ряде генов, которые должны работать согласованно для того, чтобы передне-задняя ось личинки сформировалась правильно (Klomp et al., 2015). Более того, сам принцип, использующийся для «разметки» головы и хвоста у дрозофил и у комаров-звонцов оказался, по сути, противоположным (Klomp et al., 2015). То есть, здесь различия такого масштаба, которые явно не получить единственной случайной мутацией.
Но ведь мухи и комары относятся к одному отряду насекомых (двукрылые). А формирование «переда и зада» в ходе развития личинки – это фундаментальный этап, критически влияющий на весь дальнейший ход этого развития. Так каким же образом получилось, что гены, отвечающие за формирование головы и (пардон) зада насекомого – получились разными у представителей одного и того же отряда насекомых? Как такое могло произойти, если исходить из теории эволюции?
Допустим, в далеком прошлом жил некий общий предок всех двукрылых (или вообще всех насекомых). И каждой особи этого «общего предка» на раннем этапе индивидуального развития всякий раз надо было определяться - на каком конце тела у неё будет находиться голова, а на каком конце – хвост. Для решения этой задачи, у воображаемого «общего предка всех двукрылых насекомых» должен был быть (уже) какой-то специальный биохимический механизм, который обеспечивался соответствующими генами. Это были очень важные гены. Ведь без них можно получить вообще безголовую личинку. Или, наоборот, личинку с двумя головами (где вторая голова будет вместо хвоста).
И вот получается, что в один прекрасный день, в какой-то линии потомков этого «общего предка» - гены, формирующие голову и хвост, просто взяли, и заменились на совершенно другие. На такие, которые размечают передне-заднюю ось насекомого с помощью совершенно другого принципа работы (Klomp et al., 2015). Причем, чтобы этот механизм работал, несколько новых генов должны взаимодействовать друг с другом определенным образом.
Интересно, как такое вообще могло произойти, да еще и чисто случайным путем?
Существует хорошая русская поговорка:
- Коней на переправе не меняют (С)
То есть, начинать в критический момент заменять чего-нибудь уже работающее на нечто другое – чревато не слишком удачным исходом. А ведь в нашем случае меняют даже не коней – заменяются такие гены, которые, по сути, определяют – получится ли насекомое жизнеспособным, или же оно обречено погибнуть вследствие критического уродства? Интересно, какова вероятность такой успешной «замены генов на переправе»? Еще раз подчеркнем – здесь была замена не одного (уже работающего!) гена, а нескольких. Причем замена на такие новые гены, у которых еще и подход к этой (старой) работе совершенно другой.
И такая замена должна была осуществиться не в результате продуманного генетического дизайна (что легко можно себе представить), а просто «методом случайного тыка» - серии случайных мутаций, случайно изменявших генотип насекомого в случайных местах… Тем не менее, мы наблюдаем то, что наблюдаем – у двух насекомых, принадлежащих к одному и тому же отряду, имеются совершенно разные генетические пути формирования передне-задней оси личинки.
Причем этот удивительный факт мы созерцаем на фоне… упорного сходства (других) управляющих генов у таких животных, которые разделены полумиллиардом лет (предполагаемой) эволюции – это и сходные гены (Pax), управляющие «разметкой» глаз у медуз, мух и мышей. И комплекс сходных управляющих генов, который формирует конечности позвоночным и крылья насекомым (на ранних стадиях), при этом демонстрируя целый ряд повторяющихся аналогий. Причем аналогий весьма подозрительных, поскольку конечности позвоночных и крылья насекомых, вообще-то, не гомологичны друг другу.
Что и говорить, «неисповедимы пути твои, эволюция».
Однако в контексте нашей беседы о «генетических доказательствах эволюции», нам здесь важно вот что – сегодня известны многочисленные факты, когда далекие биологические таксоны почему-то имеют сходные гены (выполняющие сходные функции)… но также известны факты, когда близкие биологические таксоны почему-то имеют разные гены, хотя эти гены задействованы в важнейших процессах индивидуального развития. Вследствие чего их изменение «на ходу» трудно себе представить.
Вот такие вот они неисповедимые, эти «генетические доказательства эволюции».
ЧТО ВСЕ ЭТО ЗНАЧИТ?
Итак, на фронте «генетических доказательств эволюции» дела у дарвинистов сегодня обстоят нехорошо. Настолько нехорошо, что хуже уже, наверное, и не надо. То есть, уже ясно, что молекулярно-генетические факты не подтверждают версию дарвиновской эволюции. Вместо стройной генетической картины, которая бы четко показывала происхождение одних живых существ из других - мы наблюдаем какую-то «генетическую кашу»: 1) массовые генетические «заимствования» между биологически удаленными таксонами; 2) в том числе, с участием фактов «генетической конвергенции» (крайне подозрительных совпадений между целыми генными комплексами у совершенно разных живых существ); 3) в то время как у родственных организмов могут наблюдаться резкие различия в генетических механизмах даже фундаментальных (жизненно-важных) процессов. Понятно, что такая картина не подтверждает версию дарвиновской эволюции, а скорее, опровергает её.
Однако возникает вопрос – тогда на что же, собственно, указывают известные сегодня молекулярно-генетические факты? О каком возможном сценарии они говорят?
Давайте попытаемся в этом разобраться – еще раз, внимательно, и с самого начала. Теперь, когда мы уже знаем: 1) что такое генетический код, ДНК и гены, 2) в чем состоит разница между значимыми нуклеотидными заменами и синонимичными, 3) какие именно возмутительные факты (со стороны молекулярной генетики) не вписываются в картину дарвиновской эволюции и почему - теперь нам уже будет проще во всем этом разобраться.
Начнем с самого начала - почему «генетические доказательства эволюции», собственно, доказывают эволюцию? Ведь что, по сути, демонстрируют молекулярно-генетические факты? Что одни группы организмов генетически ближе друг к другу, чем другие. Но такие факты – тривиальны, и ничего особо убийственного они не несут (для любых концепций, хоть эволюционных, хоть вообще не эволюционных).
Допустим, в ходе генетических исследований мы установили, что тот или иной набор генов у разных представителей крупных кошек (род Panthera) – ближе к набору аналогичных генов у мелких кошек (род Felis), чем к набору соответствующих генов у любого из представителей семейства собачьих (Canidae). Ну и что мы докажем таким образом? Ведь и без всяких генетических фактов ясно, что крупные кошки ближе к мелким кошкам, чем к собакам. Ближе и морфологически, и физиологически, и поведенчески. Так что же тогда удивительного, что они окажутся еще и генетически ближе? Ведь мы считаем, что всё разнообразие признаков организмов – как раз и записано именно в генотипах этих организмов. Поэтому открытие еще и генетической близости у тех биологических групп, которые и так близки во всех других отношениях – является просто тривиальным фактом, из которого вообще ничего не следует (никаких «доказательств»). Действительно, если бы мы создавали таксоны кошачьих и собачьих с помощью разумного дизайна, то у нас бы все равно получилось то же самое - генетические программы разных (созданных) представителей кошачьих оказались бы ближе друг к другу, чем к генетическим программам собачьих (и уж тем более, к генетическим программам каких-нибудь антилоп). Так получилось бы просто потому, что признаки, которые мы запрограммировали в генах кошачьих, отличаются от аналогичных признаков (которые мы запрограммировали в генах собачьих) в большей степени, чем эти же признаки между разными представителями кошек.
Таким образом, в самом первом приближении (из простого факта генетической близости) еще вообще ничего не следует. Такие факты можно прекрасно уложить в рамки любой концепции - хоть эволюционной, хоть не эволюционной.
Однако если мы копнем глубже и посмотрим: 1) какими именно генами могут различаться представители разных биологических групп и 2) чем именно эти гены могут различаться… то вот здесь-то как раз и всплывает эволюционная трактовка этих различий в качестве самой правдоподобной на сегодняшний день.
Речь идет о том, что у разных живых существ различаются между собой (нуклеотидными последовательностями) такие гены, которые, казалось бы, выполняют в разных организмах – одну и ту же работу. В живых клетках много такой работы, выполнять которую совершенно необходимо (практически любым организмам). Необходимо для того, чтобы живые существа вообще могли жить. Как правило, в таких (жизненно необходимых) генах закодированы белки, которые осуществляют те или иные биохимические реакции внутри живых клеток. Без этих биохимических реакций весь «конвейер» живой клетки вообще остановится.
Например, известные белки цитохромы присутствуют в клетках практически любых живых существ, участвуя там в совершенно необходимых окислительно-восстановительных реакциях. Эти химические реакции, проводимые белками-цитохромами, в принципе, одинаковы. То есть (определенные) цитохромы выполняют, по сути, одинаковую работу в клетках самых разных живых существ.
Но ведь разных живых существ в природе – несколько миллионов видов. И вот, у всех этих миллионов видов… имеется свой вариант цитохрома (!), который хотя бы чуть-чуть, но отличается (своей аминокислотной последовательностью) от цитохромов любых других (даже близких) биологических видов.
Возникает вопрос – а зачем у каждого биологического вида имеется свой собственный вариант цитохрома, если все эти цитохромы – проводят одну и ту же химическую работу?
Причем такая картина (с разнообразием вариаций, по сути, одного и того же белка) наблюдается не только с цитохромами, но и практически с любыми другими белками, имеющимися у разных биологических видов. То есть, почти бесконечная вариативность белков, выполняющих одну и ту же биохимическую работу – это общее правило, которое мы наблюдаем в живой природе.
Естественно, опять возникает тот же вопрос, но уже многократно усиленный – а какой смысл в этой бесконечной вариативности любых белков, где полный набор собственных вариантов белков имеет каждый биологических вид, если все эти вариации (аналогичных) белков выполняют, по сути, одну и ту же биохимическую работу?
Поневоле напрашивается мысль, что эти (почти бесконечные) вариации аналогичных белков – просто не имеют никакого биологического смысла. То есть, эти варианты белков – нейтральны в биологическом отношении. И любой (или почти любой) вариант такого белка – мог бы успешно выполнять в каком-нибудь «чужом» организме ту самую работу, которую он делает в своем «родном» организме. Особенно, если «чужой» и «родной» организмы являются биологически близкими видами, и если соответствующие белки различаются между собой не слишком сильно (или вообще слабо).
В общем, теоретический вывод о биологической нейтральности различий в аминокислотном составе (сходных) белков – прямо-таки напрашивается. Напрашивается вследствие тотальной распространенности в природе описанного феномена – почти бесконечного разнообразия белков, которые: 1) аналогичны друг другу по выполняемой работе; 2) при этом еще и легко узнаваемы - сходны друг с другом по многим участкам своих аминокислотных последовательностей, 3) но тем не менее, различаются у разных биологических видов (по другим участкам своих аминокислотных последовательностей или хотя бы по отдельным аминокислотам).
Более того, только что озвученный вывод (о нейтральности вариаций аминокислотного состава аналогичных белков) уже давно не является только теоретическим. Биологи на практике (во многих работах с применением генной инженерии), показали, что сходные варианты белков из разных организмов, действительно, могут заменять друг друга. И такая замена позволяет генетически модифицированному организму продолжать своё существование вполне успешно.
Например, в совсем недавней работе (Kachroo et al., 2015) биологи решили проверить, сколько аналогичных генов могут оказаться взаимозаменяемы у таких, казалось бы, далеких организмов, как человек и дрожжи. «Под нож» исследователей попали 414 генов дрожжей. В итоге, почти половину из этих генов (которые были специально удалены или просто сломаны у дрожжей) удалось заменить соответствующими человеческими вариантами этих генов. Еще точнее, дрожжи смогли эффективно задействовать человеческие варианты генов (в соответствующих биохимических путях собственных клеток) в 47% исследованных случаев (Kachroo et al., 2015).
Здесь следует еще раз подчеркнуть, что человек и дрожжи являются весьма разными организмами, в том числе, биохимически. Да и сами белки-аналоги у дрожжей и человека тоже показывают не слишком большое сходство в своих аминокислотных последовательностях. Например, для большей части аналогичных белков, исследованных в этой работе, их аминокислотные последовательности совпадали на уровне 20-50% (хотя в некоторых случаях сходство было выше или ниже этого интервала). И вот, такого уровня сходства оказалось вполне достаточно, чтобы белки, кодируемые человеческими генами, смогли эффективно работать в клетках дрожжей в половине исследованных случаев.
Что уж тогда говорить о генах между организмами более близких биологических таксонов? Их нуклеотидные последовательности могут совпадать между собой в гораздо большей степени, чем гены человека и дрожжей. Например, у человека и шимпанзе дело доходит до того, что 98% всей последовательности их нуклеотидов у целого ряда генов совершенно идентично друг другу. Естественно, напрашивается мысль, что настолько похожие гены способны заменить друг друга совсем легко.
Итак, похоже, нам следует принять тезис о биологической нейтральности наблюдаемых молекулярных различий между аналогичными генами разных организмов. Во всяком случае, согласиться с тем, что нейтральными могут быть весьма много таких различий (допустим, от 50 до 80% различий в нуклеотиотидной последовательности между аналогичными генами).
Тогда, в качестве самого простого объяснения причин существования подобных различий (между разными вариантами аналогичных генов) – можно предложить случайные нейтральные мутации. Поскольку получается, что замена значительного числа нуклеотидов в гене, кодирующем тот или иной белок, может не влиять на способность этого белка успешно выполнять свою работу, то значит, такие случайные мутации (когда они происходили) могли спокойно оставаться в геномах мутантных организмов и дальше. Ведь на приспособленность организма такие мутации не влияют. А раз так, то со временем, исходный вариант того или иного белка (его аминокислотная последовательность), в принципе, может весьма сильно измениться. Поскольку в соответствующем гене (который кодирует данный белок) будут со временем неизбежно накапливаться такие генетические изменения, которые никак не влияют на эффективность работы этого белка. Это будет происходить вследствие чисто случайных мутаций, которые тоже неизбежны.
Понятно, что такие изменения генов, из-за которых эффективность работы соответствующих белков снижалась (может быть, до нуля) – такие изменения подвергались очищающему отбору и удалялись из популяций. Ведь мутанты с такими генетическими изменениями либо вообще не имели шансов выжить, либо их шансы успешного выживания падали. Так что накапливаться со временем могут далеко не все мутации. А только такие, которые либо не влияют на приспособленность организма (нейтральные мутации), либо улучшают его приспособленность (полезные мутации). И вот, существование огромного числа разных вариантов белков-аналогов в геномах разных биологических видов - проще всего объяснить именно накоплением (со временем) в соответствующих генах нейтральных мутаций.
Но как только мы принимаем подобное объяснение причин наблюдаемых различий между генами-аналогами разных живых существ, то отсюда и начинает следовать, что молекулярно-генетические факты свидетельствуют именно об эволюции (происхождении одних организмов из других организмов). Потому что в огромном числе случаев мы наблюдаем (как я уже говорил выше), картину «постепенно убывающего сходства». Когда, например, генетические различия между представителями разных биологических видов, но в рамках одного рода (то есть, речь идет о близких биологических видах) – явно меньше, чем генетические различия между биологическими видами из разных родов. А генетические различия между представителями разных биологических родов, но в рамках одного и того же семейства, в свою очередь, тоже меньше, чем между представителями разных семейств. И так далее.
Если мы признаём тезис о нейтральности подобных различий, то получается, что разница в силе генетических отличий определяется лишь прошедшим временем, за которое эти отличия успели накопиться. Но поскольку в только что описанных случаях степень генетических различий тоже различается (между более близкими и менее близкими биологическими группами), то получается, что геномы одних живых существ существовали раздельно друг от друга в течение большего времени, чем геномы других живых существ. То есть, близкие биологические виды, принадлежащие к одному и тому же роду – существовали раздельно друг от друга в течение самого меньшего промежутка времени, в то время как более далекие биологические виды, принадлежащие к разным биологическим родам, существуют раздельно друг от друга уже больше времени. А представители разных семейств – еще больше времени.
Исходя из таких фактов (и еще, конечно, из постулата о нейтральности значительного числа молекулярных различий между генами-аналогами), теоретически имеется два возможных сценария развития жизни:
1. Либо мы должны сделать заключение об имевшей место эволюции. То есть, о строгом происхождении одних живых существ из других живых существ.
Потому что для того, чтобы объяснить разницу в степени генетических различий между разными группами организмов, нам приходится сделать вывод о наличии каких-то общих предков у этих биологических групп. Где близкие биологические группы имели некоего общего предка еще сравнительно недавно. Но потом линии этих биологических групп разошлись друг от друга (в ходе эволюции). И теперь эти (близкие) биологические группы имеют мало генетических отличий потому, что они отделились от своего общего предка недавно (соответственно, нейтральные генетические различия еще не успели накопиться). А далекие биологические группы имеют много генетических отличий друг от друга потому, что их последний общий предок жил уже давно, и поэтому после разделения этих групп в соответствующих генах-аналогах уже успело накопиться гораздо больше нейтральных различий, чем в предыдущем случае.[72]
В качестве наглядного примера рассуждений в рамках именно подобного (эволюционного) сценария, можно привести соответствующую цитату из известной сетевой версии «Доказательств эволюции»: [73]
«…Сравним теперь аминокислотные последовательности того же самого фрагмента цитохрома b у шимпанзе, человека и макаки резуса:
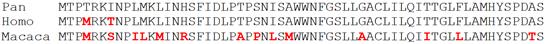
Как видим, у макаки аминокислотная последовательность этого белка сильнее отличается от человеческой и шимпанзиной, чем последовательности первых двух видов друг от друга (14 аминокислотных различий между макакой и шимпанзе, 13 - между макакой и человеком, 2 - между шимпанзе и человеком). Это полностью соответствует биологической систематике и эволюционному дереву (шимпанзе - гораздо более близкий родственник человека, чем макака)…»
Итак, логика эволюционного подхода, который в подобных фактах молекулярной генетики видит отражение именно эволюции живых существ, является вполне понятной. Даже можно сказать, очевидной. И в целом, устанавливаемые молекулярно-генетические факты подтверждают такой (эволюционный) сценарий. Как я уже говорил, картина «постепенно убывающего генетического сходства» (между всё более удаленными биологическими таксонами), в целом, действительно, наблюдается. Более того, имеет массовый характер.
Кстати, в рамках эволюционного сценария, дополнительным свидетельством в пользу нейтральности многих различий в генах и белках разных организмов может служить тот факт, что между гомологичными генами близких биологических видов чаще наблюдаются синонимичные отличия, чем значимые. Этот факт можно объяснить тем, что синонимичные отличия в генах вообще не изменяют аминокислотную последовательность белка. Следовательно, не изменяют и его химические свойства. Поэтому они всегда нейтральны в этом отношении. В то время как значимые отличия в генах отражаются на аминокислотном составе белка, следовательно, могут влиять на его биохимические способности. Поэтому значимые замены нейтральны далеко не во всех случаях - большая часть таких мутаций будет ухудшать работу соответствующего белка. Такие мутации имеют низкую вероятность закрепиться в генотипе. Вот поэтому в генах близких видов мы обычно и наблюдаем меньше значимых замен, чем синонимичных. То есть, и этот наблюдаемый факт (преобладания синонимичных замен над значимыми) тоже хорошо объясняется в рамках именно эволюционного сценария (постепенного накопления мутаций).
Наконец, в пользу эволюционного сценария говорит и тот факт, что даже внутри биологических видов (между разными популяциями и даже между разными особями одного вида) тоже наблюдаются молекулярно-генетические различия. Пусть и малые. Этот факт как бы намекает на то, что с момента происхождения данного биологического вида (от какой-то исходной предковой популяции) прошло еще мало времени. Но за это время между разными популяциями этого вида уже успели накопиться кое-какие мутации. То есть, молекулярно-генетическое разнообразие, наблюдаемое внутри биологических видов – в русле эволюционного подхода выглядит естественным продолжением (или наоборот, началом) тех самых эволюционных процессов, которые уже привели к появлению разных биологических видов, родов, семейств и так далее (за гораздо больший промежуток времени).
2. Однако чисто эволюционная картина начинает выглядеть совсем не так хорошо, как хотелось бы, стоит нам только вспомнить о многочисленных филогенетических конфликтах, о которых мы говорили чуть выше. Молекулярно-генетические исследования показали, что феномен «постепенно убывающего генетического сходства» (указывающий на эволюционный сценарий), сохраняется в целом для большинства биологических групп, но тем не менее, часто нарушается, в самых разных биологических таксонах (филогенетические конфликты). Причем целый ряд таких филогенетических конфликтов не может быть объяснен даже «горизонтальным переносом генов». Потому что в этих случаях генетические заимствования имеют слишком массированный характер (см. выше). К сегодняшнему дню число обнаруженных «филогенетических конфликтов» так размножилось, что некоторые исследователи уже прямо говорят – взаимоотношения биологических групп не выглядят, как эволюционное дерево.[74]
И такие факты нельзя просто игнорировать. Потому что в рамках эволюционного подхода, у конкретного биологического вида обязательно должен быть какой-то конкретный эволюционный предок. Не может быть так, чтобы один комплекс (из сотен) генов в генотипе какого-нибудь биологического вида – брал своё начало от одного общего предка, а другой генетический комплекс в генотипе этого же вида – брал своё начало уже от совершенно другого «общего предка».
Если же такая картина всё-таки наблюдается (по факту), то это обстоятельство сразу же и подрывает эволюционный сценарий происхождения данного биологического таксона. Ну а если таких «генетических парадоксов» уже установлено много, и они оказываются «рассыпаны» по всему живому царству - тогда чисто эволюционный сценарий происхождения живых существ подрывается уже в целом.
Таким образом, на сегодняшний день, на основании именно молекулярно-генетических фактов, можно заключить, что развитие жизни на Земле часто шло в соответствии с эволюционным сценарием, но этот сценарий регулярно нарушался. То есть, картина получается сложнее, чем это ожидается в рамках современного дарвинизма.
Возникает вопрос – какая же сила могла регулярно нарушать эволюционный сценарий происхождения организмов? Очевидно, что никакие естественные природные силы не могут «лепить» генотип конкретного живого существа из разных генотипов неродственных организмов. Такие вещи (горизонтальный перенос генов) наблюдаются у прокариот (бактерии и архебактерии). Но, во-первых, наблюдаются в очень ограниченном объеме – с помощью горизонтального переноса «перескакивают» от одной бактерии к другой одиночные гены (или в крайнем случае, можно предположить перенос группы генов). А во-вторых, подобные вещи наблюдаются только у прокариот. А у эукариот - горизонтальные переносы генов пока, вроде бы, никто (напрямую) не наблюдал. И это не случайно – успешный горизонтальный перенос гена в случае многоклеточного эукариота осуществить гораздо сложнее, чем в случае одноклеточной бактерии. Более того, многие бактерии имеют специальные механизмы для захвата чужих генов (в отличие от эукариот).
Тем не менее, мы видим (по факту), что у некоторых эукариотических организмов целые генетические комплексы, состоящие из сотен генов, и отвечающие за формирование сложного биологического признака – могут оказаться сходными с аналогичными генетическими комплексами организмов из таких биологических групп, от которых они произойти никак не могли. Очевидно, что в этом случае все возможные естественные гипотезы уже заканчиваются - мы понимаем, что ни одна естественная природная сила не могла приводить к столь массированным аналогиям в генетических механизмах, обеспечивающих формирование и работу сложных органов у совершенно разных живых существ. И нам остаётся только вводить в сценарий развития жизни какого-то разумного «генного инженера».
Например, можно предположить, что в истории Земли разные группы организмов действительно возникали в разное время, но не путем их эволюции друг из друга, а вследствие их независимого создания в разное время (одни организмы позже, другие раньше). В этом случае мы бы тоже наблюдали примерно похожую картину – между генами-аналогами живых существ тоже наблюдалась бы разная степень накопленных генетических отличий – биологические группы, созданные раньше, накопили бы больше генетических различий, чем группы, созданные позже.
И теперь, если мы сделаем еще одно дополнительное предположение – что более поздние биологические таксоны могли создаваться не обязательно независимо, а может быть, на основе уже имеющихся генотипов (тех биологических таксонов, которые были созданы раньше), и при этом, для создания некоторых сложных признаков в новых биологических таксонах могли задействоваться те инженерные решения, которые ранее уже были применены при создании совершенно других биологических групп… Тогда мы получаем именно такую картину развития жизни на Земле, в пользу которой говорит вообще весь комплекс наблюдаемых молекулярно-генетических фактов (а не только их выборочный набор).
Более того, получают свое объяснение и такие факты, когда, казалось бы, родственные организмы, тем не менее, имеют радикальные отличия в генетических механизмах даже фундаментальных (жизненно-важных) процессов развития (см. выше пример с мухами и комарами). Действительно, той разумной силе (которая предполагаемо создавала разных живых существ) - никто ведь, наверное, не запрещал создавать какие-то биологические таксоны с использованием разных инженерных решений?
Таким образом, на сегодняшний день, генетические факты больше всего свидетельствуют в пользу развития жизни по тому сценарию, который предлагает концепция непрерывного творения. А вот в пользу современного дарвинизма (т.е. в пользу строго естественной эволюции) можно привести одни генетические факты, но другие генетические факты – либо недостаточно гладко укладываются в эту концепцию, либо вообще в неё не укладываются.
Во второй части книги мы проведем подробный анализ концепции непрерывного творения в сравнении с современным дарвинизмом в свете всего комплекса установленных сегодня биологических фактов (не только молекулярно-генетических).
Но сейчас нам надо озвучить еще один возможный (третий) вариант, который тоже может вытекать из известных сегодня молекулярно-генетических фактов, но который мы пока еще не рассматривали.
3. Дело в том, что все рассуждения, которые мы только что привели выше, справедливы только в том случае, если мы принимаем постулат о полной нейтральности разных вариантов аналогичных белков, наблюдаемых у разных организмов. Но насколько является доказанным этот теоретический постулат на практике?
Выше я уже приводил пример, когда человеческие варианты генов смогли успешно заменить соответствующие варианты генов дрожжей примерно в половине всех исследованных случаев. Но насколько успешно они их заменили?
Этот вопрос далеко не праздный. Например, сегодня людям, страдающим дефектами клапанов сердца, могут проводить соответствующие операции с протезированием клапана. То есть, с заменой их собственного клапана на искусственный аналог. И люди после таких операций продолжают жить, причем во многих случаях – весьма долго. Но вряд ли кто-нибудь из нас будет оспаривать тот факт, что если собственный (естественный) сердечный клапан является полностью здоровым, то он все-таки лучше протеза?
Так же и здесь. Да, человеческие варианты генов смогли успешно заменить гены дрожжей. Но от такой замены приспособленность дрожжей (с «имплантированными» генами) – точно ли не изменилась? А если эта приспособленность снизилась на 20%? Да, дрожжи продолжают жить, но делают это хуже. В этом случае разве сможем мы сказать, что разница в нуклеотидных последовательностях генов-аналогов дрожжей и человека – биологически нейтральна?
Нет, мы так сказать уже не сможем (это будет не так по факту). Но тогда и объяснение разницы в нуклеотидных последовательностях этих генов-аналогов – только временем, прошедшим со дня разделения этих биологических групп - уже будет звучать не так убедительно, как раньше. Потому что, может быть, эти гены-аналоги различаются между собой не просто так, а именно для того, чтобы человеческий вариант этого гена работал максимально эффективно в тех биохимических условиях, которые предоставляет именно организм человека. А «дрожжевой» вариант этого гена имеет такие особенности для того, чтобы самым лучшим образом (из всех возможных) работать именно в клетках дрожжей?
В общем, в этом случае, наши «генетические доказательства эволюции» сразу исчезают. Потому что эти доказательства основаны на предположении о нейтральности наблюдаемых различий в генах-аналогах разных организмов. Но если соответствующие различия, на самом деле, не нейтральны, тогда от «генетических доказательств эволюции» мало что остаётся.
То же самое можно сказать и о близких биологических таксонах. Выше мы привели конкретный пример, где разные варианты цитохрома b (человека, шимпанзе и макаки) различались у человека и шимпанзе меньше, чем оба эти варианта – от варианта гена макаки. Однако отличия во всех трех случаях были небольшими. А в случае человека и шимпанзе – совсем небольшими. Мы этот факт интерпретировали в рамках эволюционного сценария. Тем более, если учесть тот факт, что на Земле миллионы биологических видов, и почти у каждого такого вида – имеется уже свой вариант цитохрома. Очевидно, что самым правдоподобным объяснением такого разнообразия вариантов цитохромов b (даже у близких биологических видов) – является постулат о биологической нейтральности подобных вариаций.
Но что, если мы ошибаемся? Если это потрясающее разнообразие разных вариантов генов, которое мы наблюдаем в живой природе – на самом деле, не является нейтральным? А представляет собой, допустим, «тонкую настройку» каждого биологического вида? Такую тонкую настройку, которая необходима именно для того, чтобы данный биологический вид был именно этим биологическим видом, а не каким-нибудь другим? Допустим, каждый собственный вариант гена является наиболее оптимальным именно для своего биологического вида. Но эта оптимальность может выражаться в совсем низких величинах, например, в долях процента. Тогда, по всей совокупности генов (каждый из которых уникален для своего биологического вида) – в совокупности будут «набегать» уже многие проценты дополнительной приспособленности. Которые, может быть, и обеспечивают биологическому виду ту самую устойчивость и индивидуальность, которую биологические виды нередко демонстрируют (см. ниже, во второй части книги).
Понятно, что такое предположение, о настолько тщательно продуманном генетическом дизайне каждого биологического вида - выглядит очень спекулятивно. Так, будто мы стараемся спастись любыми путями от очевидного вывода о биологической нейтральности разных генетических вариаций генов-аналогов у разных видов. Действительно, трудно поверить, что нуклеотидные последовательности разных биологических видов могут быть настроены столь тонко (вплоть до синонимичных различий между видами).
И тем не менее. Наука уже давно привыкла к тому, что такие фразы как «очевидный вывод» и «трудно поверить» - на самом деле вообще ничего не доказывают (несмотря на всю их «очевидность»). В истории науки было немало примеров, когда «совершенно очевидные вещи» со временем переходили в разряд ошибочных, в то время как гипотезы, которым было «трудно поверить» - оказывались правильными, встраивались в систему научных знаний и со временем становились привычными. Поэтому реальными доказательствами в науке являются не «очевидные выводы о наибольшей правдоподобности» - а результаты конкретных эмпирических исследований. Которые и показывают, действительно ли то, что кажется очевидным, соответствует тому, что имеется в реальной природе.
Таким образом, наши «очевидные выводы» о нейтральности различий между разными вариантами генов-аналогов у разных организмов – необходимо подтверждать результатами соответствующих исследований. Но как это сделать на практике?
Допустим, замена человеческого варианта гена на ген макаки (в озвученном выше примере), и правда, не является биологически нейтральной. А приведет к тому, что у человека с «имплантированным» цитохромом макаки эффективность проведения соответствующей химической реакции в клетке снизится на 1%. Однако разных генов у человека много, и все они, так или иначе, тоже влияют на общую приспособленность. В результате, замена только одного обсуждаемого цитохрома, возможно, снизит общую (итоговую) приспособленность человека даже не на один процент, а вообще на 0.01%.[75] Как же в этом случае реально проверить, нейтральны ли наблюдаемые молекулярные различия между генами, или же они не случайны? Ведь разницу в 0.01% приспособленности просто нельзя уловить на практике не только в экспериментах с разными видами приматов, но даже в экспериментах с разными бактериальными колониями. То есть, вряд ли можно уловить разницу (на уровне статистической значимости) в скорости роста разных колоний бактерий в пределах 0.01%.
Ну а если цитохром шимпанзе (имеющий вообще мало отличий от цитохрома человека) – при «имплантации» снизит эффективность соответствующей биохимической реакции у человека не на 1%, а вообще на 0.1%? И тогда разница в итоговой приспособленности может составить вообще 0.001%. Как в этом случае проверить, нейтральны ли те несколько различий между нуклеотидами, которые имеются в цитохромах шимпанзе и человека, или же эти различия всё-таки «тонко настроены» (под каждый вид)?
Тем более сложно будет это сделать, если в наших теоретических рассуждениях (о возможности «тонкой генетической настройки» организмов) мы допустим более мягкий, компромиссный вариант. Допустим, большинство нуклеотидных различий между генами-аналогами, действительно, имеют не случайный характер (являются «тонкой настройкой»), однако малая часть таких различий – все-таки может быть результатом чисто случайных мутаций (которые успели закрепиться за время существования данного биологического таксона). В этом случае мы получим еще более туманную картину, которую проверить будет еще труднее.
И наконец - что если какая-то часть генов (в рамках конкретного биологического вида) специально предусмотрена быть особенно изменчивой (именно для этого вида)? Чтобы этот вид, допустим, имел возможность приспособиться к разным условиям среды? Или просто в целях разнообразия. Например, повышенное разнообразие цвета волос, глаз и кожи у человека (или черт его лица) – возможно, планировалось изначально? Действительно, почему у людей в избытке наблюдаются брюнеты, шатены и блондины (да еще и самых разных «оттенков»)? Ведь возраст современных рас человека (не говоря уже о разных народностях) не должен превышать нескольких десятков тысяч лет.
То есть, какая-то часть генетической изменчивости может быть вообще «заранее запланированной» (о чем может свидетельствовать явно повышенная частота мутаций именно по этим локусам). Еще какая-то часть наблюдаемых различий может быть тоже нейтральной, но эти мутации не «планировались заранее», а действительно, являлись чисто случайными и просто накопились за время существования этого биологического вида. И наконец, еще какая-то часть различий может быть слабо-вредными (или слабо-слабо-вредными) мутациями, которые тоже успели накопиться за время существования этого биологического вида. Такие мутации постепенно ломают (искажают, разрушают) ту изначально тонкую генетическую настройку (тот уникальный набор вариантов генов, идеально подходивших именно этому биологическому виду), который имелся у данного вида изначально (с момента создания). Как теперь строго проверить, какие из озвученных теоретических возможностей действительно имеют место (в наблюдаемом генетическом разнообразии), и если имеют, то насколько значимую долю составляет каждая из них?
Похоже, что строго проверить постулат об абсолютной нейтральности (так же как и об абсолютной случайности) «тонких нуклеотидных различий» сегодня вряд ли возможно.
А вот обратные факты, то есть такие, которые могли бы свидетельствовать в пользу наличия у подобных замен определенного биологического смысла – вполне можно попытаться обнаружить. И выше я уже приводил такие исследования. Действительно, уже имеется целый ряд исследований, результаты которых свидетельствуют в пользу того, что определенный биологический смысл могут иметь даже синонимичные замены (не говоря уже о значимых). Еще раз озвучим некоторые из этих исследований. В работе (Xu et al., 2012) было показано, что разница между синонимичными заменами может приводить к изменению биологических часов у цианобактерий. В другой работе было показано, что синонимичные замены могут приводить к аналогичным последствиям (сбою биологических часов) у плесени (Zhou et al., 2012). В работе (Cuevas et al., 2002) тоже было показано, что многие синонимичные замены могут иметь биологический смысл. Поскольку в разных линиях вирусов, независимо подвергавшихся одним и тем же вызовам среды на протяжении ряда поколений, в итоге возникла повторяемость одних и тех же мутаций, в том числе, и синонимичных замен. Что указывает на какую-то биологическую роль этих замен. Основываясь на полученных результатах, авторы спекулятивно подсчитали, что действительно нейтральными (у исследованных ими вирусов) можно считать только долю процента (0.28%) всех возможных мутаций (Cuevas et al., 2002).[76] Кроме того, авторы сослались еще на одну работу, где была проведена примерная оценка доли нейтральных мутаций у человека. Эта оценка составила менее 20% (Fay et al., 2001).[77]
Последняя цифра не слишком стыкуется с предположением о нейтральности миллионов вариантов нуклеотидных различий в генах-аналогах тысяч биологических видов. Где степень различия между белками-аналогами может достигать 85% (у далеких биологических групп). А оценка 0.28% нейтральных мутаций, полученная авторами работы с вирусами (Cuevas et al., 2002) - вообще не стыкуется с представлениями о нейтральности хотя бы заметной части нуклеотидных замен.
Понятно (еще раз), что если результаты, полученные в озвученных работах, имеют широкое распространение в природе, то есть, если по-настоящему нейтральных замен, на самом деле, мало, а большая часть различий нуклеотидной последовательности в генах-аналогах разных биологических видов всё-таки имеет какое-то значение для приспособленности этих видов (представляя собой очень тонкую генетическую настройку) - тогда от «генетических доказательств эволюции» практически ничего не остаётся. Потому что в этом случае молекулярно-генетические факты не будут противоречить тезису о независимом (раздельном) создании даже биологических видов (в рамках одного рода). Не говоря уже о биологических таксонах более высокого ранга.
Однако вполне может быть наоборот - что результаты только что озвученных исследований представляют собой довольно редкое исключение из правил (когда даже синонимичные различия имеют определенное биологическое значение, например, для механизма биологических часов). Но в целом, значительная часть нуклеотидных различий между генами-аналогами самых разных организмов, всё-таки, нейтральна.
В этом случае (еще раз), наиболее правдоподобным становится сценарий непрерывного (постепенного) творения одних видов на основе других (по крайней мере, в значительном числе случаев).
Какой из этих двух сценариев больше соответствует действительности, наверное, должны показать будущие исследования.
Ну а закончить наши глубокие размышления (о том, что же доказывают «генетические доказательства» и доказывают ли вообще что-нибудь) я предлагаю на бодрой юмористической ноте. Для этого давайте еще раз обратим внимание на ту пикантную особенность, которой «генетические доказательства эволюции» уже успели прямо-таки прославиться. А именно, полюбуемся на их «умопомрачительную точность».
Еще раз вернемся к тому самому роду пантер (Panthera), о котором мы уже говорили выше - там, где снежный барс неожиданно «примкнул» к тигру, льву, леопарду и ягуару. И просто посмотрим, что же показали разные молекулярно-генетические исследования в отношении происхождения этих замечательных зверей.
Для этого открываем соответствующую работу (Davis et al., 2010) и смотрим на соответствующий рисунок (24):
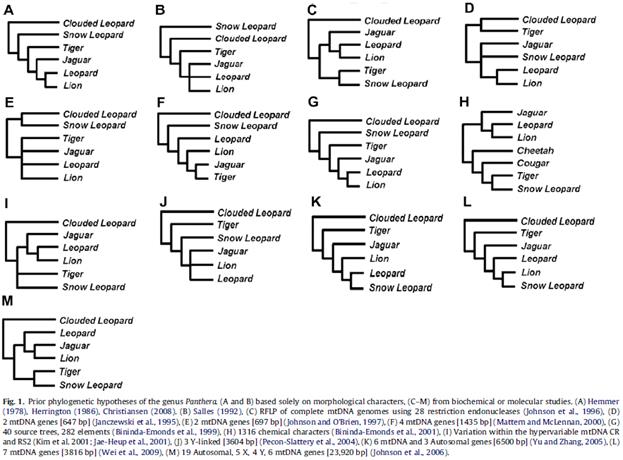
Рисунок 24. Возможные варианты происхождения разных видов рода Panthera и некоторых «сопредельных» родов кошачьих, предложенные разными (независимыми) исследованиями. На рисунке: Clouded Leopard – дымчатый леопард (Neofelis nebulosa); Snow Leopard – снежный барс ( Uncia uncia ); Tiger – тигр (Panthera tigris); Jaguar – ягуар (Panthera onca); Leopard – леопард (Panthera pardus ); Lion – лев (Panthera leo); Cheetah – гепард ( Acinonyx jubatus ); Cougar – пума (Puma concolor ). Рисунок взят из работы (Davis et al., 2010). Внизу сохранены оригинальные ссылки на те исследования, результаты которых привели к построению того или иного варианта филогении.
На этом рисунке показаны разные варианты эволюционного происхождения крупных кошачьих, полученные на основании результатов разных исследований. Итак:
Варианты А и B построены на основе морфологических признаков. В обоих вариантах наиболее удаленными от всех представителей рода пантер оказываются дымчатый леопард (Neofelis) и снежный барс (Uncia). Собственно поэтому этих животных и выделили (в свое время) в отдельные роды. Потому что морфологически данные виды, действительно, гораздо сильней отличаются от любого из представителей рода пантер, чем виды рода Panthera друг от друга.
Различия между вариантами А и В состоят в том, что в одной классификации (А) снежный барс оказывается ближе к пантерам, чем дымчатый леопард, а в другой классификации (В) наоборот. И еще в классификации (А) наиболее близкими друг другу показаны лев и леопард, затем ягуар, а потом только тигр. А в классификации (В) лев, леопард и ягуар показаны на одинаковом расстоянии друг от друга.
А вот дальше идут результаты, полученные в разных биохимических и молекулярно-генетических исследованиях:
Вариант С – тигр вдруг (!) оказывается более близким к снежному барсу, чем к любым другим представителям своего рода (Panthera).
Вариант D – относительно близки друг другу оказываются уже тигр и дымчатый леопард. В то время как сам тигр опять «выпадает» из рода Panthera (оказывается наиболее удаленным от них вместе с дымчатым леопардом). Напротив, снежный барс оказывается более близок к оставшимся представителям рода Panthera, причем на таком же уровне близости, как и ягуар (лев и леопард остаются наиболее близки между собой).
Вариант Е – дымчатый леопард и снежный барс оказываются более близки друг к другу, одновременно максимально удаляясь от всех других представителей рода Panthera. В то время как разные представители рода пантера оказываются равноудаленными друг от друга (включая тигра).
Вариант F – наиболее близкими здесь оказываются уже тигр и ягуар (совсем близки)! К этой парочке вплотную примыкает лев. А леопард становится наиболее отодвинутым от озвученной троицы.
Вариант G – полностью повторяет вариант (А) морфологической классификации.
Вариант H – здесь уже добавляются новые действующие персонажи. Однако лев и леопард опять оказываются наиболее близки друг другу. И к ним (опять) довольно тесно примыкает ягуар. В то же время, тигр снова оказывается максимально удален от них (от льва, леопарда и ягуара), и опять в качестве ближайшей родни выбирает себе снежного барса. Более того, к ним теперь еще и приближается пума. В то время как гепард выглядит довольно удаленным (и уединенным). Но все же он оказывается ближе к тигру, чем лев, леопард и ягуар.
Вариант I – лев и леопард наиболее близки, к ним примыкает ягуар. Тигр и снежный барс равноудалены от этой троицы, но при этом не слишком близки и друг к другу. Самый удаленный – дымчатый леопард.
Вариант J – наиболее удален от всех других видов дымчатый леопард. За ним следует (по степени отрыва) тигр. А снежный барс оказывается ближе к «троице»: ягуар, лев, леопард. Которые, в свою очередь, оказываются равноудаленными друг от друга (но при этом более близки друг другу, чем снежный барс).
Вариант K – вот так сюрприз! Наиболее близкими друг другу оказываются снежный барс и леопард (до этого такое сочетание еще не встречалось). К ним вплотную примыкает лев, затем ягуар. Ну а потом идет тигр и уже дымчатый леопард.
Вариант L – снова новое сочетание! Ближайшими генетическими родственниками оказываются снежный барс и лев! Затем с небольшим отрывом следует леопард. За ним ягуар. За ним тигр. И наконец, дымчатый леопард.
И наконец, вариант M – ближайшими генетическими родственниками снова оказываются тигр и снежный барс. При этом данная парочка видов сравнительно удалена от троицы, состоящей из леопарда, ягуара и льва. Причем в рамках этой троицы тоже возникает новое сочетание – лев и ягуар оказываются наиболее близки друг к другу, в то время как леопард оказывается удаленным от этих видов в несколько большей степени (а наиболее удаленным оказывается опять дымчатый леопард).
Вот такой вот «винегрет». Комментируя подобные «результаты молекулярно-генетических исследований», авторы данной работы написали (Davis et al., 2010):
«Несмотря на многочисленные публикации на эту тему, нет двух молекулярных исследований, которые бы реконструировали Panthera с той же самой топологией»
И видимо, в качестве выхода из этой ситуации, авторы работы (Davis et al., 2010) предложили… уже свой вариант филогении обсуждаемых видов крупных кошек. Естественно, тоже на основании соответствующих молекулярных исследований, причем весьма обширных. В этом варианте классификации лев и леопард опять становятся наиболее близкими друг другу. К этим видам уже примыкает ягуар. А наиболее удаленными от этой троицы снова оказываются тигр и снежный барс. Которые, в свою очередь, не слишком близки друг к другу, но всё же ближе, чем к льву, леопарду и ягуару.
Вот на этой окончательной ноте я бы и хотел закончить…
Если бы в 2011 году не вышла новая (очередная) работа (Wei L. et al., 2011), в которой лев и снежный барс снова оказались настолько близки друг к другу, что даже потеснили в этом деле леопарда (леопард оказался более удален от льва, чем лев от снежного барса).
Ну а в 2013 году… (правильно!) вышла новая работа, которая утверждает, что опять «восстановила справедливость», указав на льва и леопарда, как на наиболее близкие виды, и немного отодвинув от этой парочки снежного барса (Bagatharia et al., 2013). То есть, снежный барс опять оказался близким родственником льву, но все же менее близким, чем леопард.
Похоже, это может продолжаться бесконечно – непонятно, сколько еще времени молекулярно-генетические исследования будут «радовать» нас бесконечным тасованием своих «филогенетических колод». И в зависимости от того, какие конкретно массивы разных генов были взяты для анализа – знакомить с новыми и новыми вариантами «эволюционных путей»… Интересно, если бы мы так же почтительно слушали профессиональных гадателей на кофейной гуще, которые, пристально вглядываясь «во глубину кофейных гущ», черпали бы оттуда откровения о возможных путях эволюции крупных кошек… мы бы провели время с такой же пользой?
Дата: 2019-02-02, просмотров: 547.