Начиная с 1940-х годов два микробиолога, которые бежали в США из Европы, оказавшейся под властью Адольфа Гитлера, — итальянец Сальвадор Лурия (Salvador Luria, 1912–91) и американец немецкого происхождения Макс Дельбрюк (Max Delbrück, 1906–81) — разработали важнейшую методику, обогатившую генетические исследования. Они изучали свойства группы вирусов-бактериофагов («пожирателей бактерий»). Любая из известных бактерий является добычей для хотя бы одного из этих вирусов, которые состоят из ДНК, окруженной белковой оболочкой. Бактериофагов легко хранить в лаборатории, а их действие на клетку-хозяина поистине поражает воображение — за какие-то несколько минут после заражения бактерия-хозяин оказывается взломанной и производит на свет не меньше сотни идентичных копий исходного вируса. Очевидно, что что-то в вирусе передает генетическую информацию потомкам, но что — белки или ДНК?
Ответ на этот вопрос дал эксперимент Херши и Чейз. Методика, использованная Алфредом Херши и его коллегой Мартой Коулз Чейз , проста в описании. Они выращивали две группы бактерий: одну в среде, содержащей радиоактивный фосфор-32, другую — в среде с радиоактивной серой-35. Бактериофаги, добавленные в среду с бактериями и атаковавшие их, поглощали эти радиоактивные маркеры. Чтобы понять происходившие далее события, надо знать, что фосфор входит в состав ДНК (он находится в фосфатных группах в ядре), но отсутствует в белковой оболочке вируса. Сера же, наоборот, входит в состав белка, но отсутствует в ДНК. Таким образом, пара радиоактивных маркеров позволяла разграничить роли двух компонентов вируса в его репродукции.
После этого ученые «натравливали» на бактерии две группы вирусов — с меченой ДНК и меченым белком. Не дожидаясь завершения процесса инфицирования, бактерии отделяли от остального материала с помощью центрифуги, а затем выявляли присутствие радиоактивной метки. Результаты говорили сами за себя: в бактериях был обнаружен фосфор-32 , а сера-35 оставалась в среде. Поскольку размножение вирусов происходит внутри бактерий, куда белки не проникают, было ясно, что это размножение может быть обусловлено только ДНК.
Сегодня нам известен механизм этого процесса: вирус прикрепляется к бактерии и вводит в нее вирусную ДНК, оставляя снаружи белковую оболочку. Вирусная ДНК встраивается в бактериальную ДНК и «переключает» генетический аппарат бактерии, заставляя его работать на себя для создания многочисленных копий вируса. После того как ресурсы бактерии будет исчерпаны, клетка разрушается, высвобождая новое поколение «штампованных» вирусов. Эксперимент Херши—Чейз со всей очевидностью показал, что гены размещены в молекуле ДНК, и это главный принцип современной науки.
Правила Чаргаффа — система эмпирически установленных правил, описывающих количественные соотношения между различными типами азотистых оснований в ДНК. Правила были сформулированы в результате работы группы биохимика Эрвина Чаргаффа в 1949-1951 годах.
К работам группы Чаргаффа царила так называемая «тетрануклеотидна» теория, согласно которой считалось, что ДНК состоит из повторяющихся блоков по четыре разных азотистых основания (аденин, тимин, гуанин и цитозин) в каждом. Чаргаффу с сотрудниками удалось разделить нуклеотиды ДНК с помощью бумажной хроматографии и определить точные количественные соотношения нуклеотидов разных типов. Они значительно отличались от эквимолярных, которых можно было бы ожидать, если бы все четыре типа были представлены в равных пропорциях. Соотношение, обнаруженные Чаргафф для аденина (А), тимина (Т), гуанин (Г) и цитозина (Ц), оказались следующими:
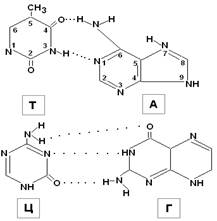
-Содержание аденина равной содержания тимина, а содержание гуанина — количеству цитозина: А = Т, Г = Ц.
-Количество пуринов равно количеству пиримидинов: А + Г = Т + Ц.
-Количество оснований с 6 аминогрупп равно количеству оснований с 6 кетогруппы: А + Ц = Г + Т (Это правило следует из первого). Вместе с тем, соотношение доля Г + Ц (содержание ГЦ) может быть разным в ДНК разных видов. В одних преобладают пары АО, в других — ГЦ.
Правила Чаргаффа вместе с данными рентгеноструктурного анализа, сыграли решающую роль в расшифровке структуры ДНК Джеймсом Уотсоном и Фрэнсисом Криком.
На основании этих правил и большого числа разнообразных экспериментальных данных в 1953 г. Д. Уотсон и Ф. Крик разработали модель ВС ДНК.
Молекула ДНК построена из двух полинуклеотидных. цепей, ориентированных антипараллельно (3′ конец одной цепи соседствует с 5′ концом второй цепи); цепи на всем протяжении связаны Н-связями. При этом каждый мононуклеотид участвует в образовании Н-связей.
Н-связи образованы между А одной цепи и Т другой, между Г одной цепи и Ц другой. Основания, образующие такие пары комплементарны друг другу, то есть между ними легче образуется Н-связь.
ПС одной цепи ДНК комплементарна ПС второй цепи, что не означает их идентичности
Обе цепи закручены вправо в спираль, имеющую общую ось. Цепи могут быть разделены только раскручиванием. Такие спирали называются плектонемическими.
Пурины и пиримидиновые основания обращены внутрь спирали. Их плоскости перпендикулярны оси спирали и параллельны друг другу, таким образом получается стопка оснований. Пентозофосфатные остовы обеих цепей обращены наружу от спирали.
Стабилизация ВС ДНК обеспечена за счет:
1) особого вида вандервальсовых сил между АЩ – так называемых стэкинг-взаимодействий;
2) водного окружения фосфатов и особенно нейтрализации их зарядов. –q стремятся оттолкнуться друг от друга, вытягивают цепь ДНК и тем самым стабилизируют ее;
3) образование двойной спирали ДНК возможно только при условии связывания фосфатов (–q) с гистонами – основными белками (+q);
4) Н-связями.
Комплекс белков с ядерной ДНК клеток называется хроматином.
Двойная спираль (-форма) Уотсона и Крика имеет размерность:
1) -спирали – 2,1 нм;
2) один виток (шаг спирали) составляет 10 пар АО; 3,4 нм;
3) высота каждого мононуклеотида 0,34 нм.
В природной ДНК хромосом строгая двуспиральность нарушается на тех участках, которые называются палиндромами (перевертышами). В таких фрагментах ДНК последовательность мононулеотидов одинакова слева направо и справа налево. Комплементарные основания палиндромов спариваются и образуют «кресты» или «шпильки». Они помогают регуляторным белкам узнавать места списывания генетического текста ДНК хромосом.
Третичная структура ДНК образуется в результате дополнительного скручивания в пространстве в виде суперспирали или изогнутой, то есть сломанной двойной спирали.
В составе хроматина ДНК сост 30-45%; гистоны 30-50%; негистиновый белок 4-33%; РНК 1,5-10%.
В электронном микроскопе хроматин напоминает бусы – шаровидные утолщения – нуклеосомы, с перемычками. Каждая нуклеосома содержит 8 молекул гистонов и намотанный на них участок ДНК, содержащий 146 пар АО. При такой укладке длина молекулы ДНК уменьшается ~ в 7 раз. Это лишь первый уровень укладки цепи. Длина молекул ДНК 3-5 см, а длина хромосом всего несколько мкм, то есть степень укорочения ДНК достигает нескольких тысяч. Это происходит в результате дополнительного скручивания нуклеосомной нитки бус.
Продолжение 8 вопроса
Репликация – это процесс удвоения ДНК, в процессе репликации каждая цепь материнской ДНК служит матрицей длясинтезы новой дочерней цепи. Репликация происходит в S-фазу клеточного цикла. Хромосома содержит 1ну двухцепочечную молекулу ДНК.
Основы для репликации:
1. Принцип комплементарности (взаимосоответствия) – способность формировать водородные связи.
2. Матричный принцип – каждая из 2х нитей – матрица для создания комплементарной дочерней цепи.
3. Полуконсервативный механизм – при удвоении ДНК новая двухцепочечная молекула содержит 1ну исходную цепь и 1ну вновь синтезированную.
Условия репликации:
1) Матрица – 2 расплетенные нити ДНК, образующие репликационную вилку.
2) Субстраты и источники энергии для синтеза – дНТФ (дАТФ, дГТФ, дЦТФ, дТТФ)
3) Ионы Mg
4) Репликативный комплекс ферментов:
ДНК-хеликаза– расплетает двойную спираль ДНК и разрывает водородные связи.
ДНК-топоизомераза 1и2 –устраняют суперспирализацию ДНК в точке разрыва, облегчая формирование репликационной вилки.
SSB-белки –связываются с одноцепочечными нитями ДНК и препятствуют их повторному комплементарному соединению.
ДНК-полимераза – катализирует образование фосфодиэфирных связей между нуклеотидами и удлинение уже существующей цепи. 2 свободных нуклеотида она соединять не может.
Праймаза– катализирует синтез олигорибонуклеотидов – коротких молекул РНК.
ДНК-лигаза – соединяет фрагменты ДНК.
Теломераза– обеспечивает построение концов ДНК.
Этапы репликации:
Инициация – формирование репликативной вилки при участии ДНК-хеликазы и ДНК-топоизомераз, в итоге происходит разделение цепей ДНК, а приссоединенные SSB-белки препятствуют повторному соединению цепей. Инициация происходит в нескольких точках – точках инициации или ориджины репликации. Единица репликации эукариот - репликон – участок между соседними ориджинами.
Элонгация – синтез новой цепи ДНК, она начинается праймазой, которая синтезирует олигорибонуклеотиды – праймеры. Далее синтез дочерней цепи ДНК происходит за счет полимеразы от 5` к 3` концу. Новые цепи синтезируются неодинаково. На 1ой цепи ДНК 5`- 3`цепь растет непрерывно, быстро и называется лидирующей. На другой нити ДНК новая цепь синтезируется в виде фрагментов Оказаки – эта цепь называется отстающей.
Терминация – удаление праймеров и завершение формирования отстающей цепи ДНК. Праймеры удаляются эндонуклеазой, а на их месте ДНК-полимераза достраивает ДНК. Сшивание фрагментов происходит за счет ДНК-лигазы. В итоге на каждой матричной ДНК формируется 2 дочернии копии ДНК.
Дата: 2019-02-02, просмотров: 381.