Ответ на 4 вопрос
Трансформацией называется перенос в определённых условиях любой части наследственной молекулы одного организма в наследственную молекулу другого организма. Другими словами — это переход отрезка ДНК одной клетки в геном другой клетки в функционально активном состоянии.
Процесс трансформации представляет собой естественный процесс, который приводит к изменению наследственности клеток.
Изучение процесса трансформации послужило толчком для открытия трансдукции — процесса перенос генов из одной бактериальной клетки в другую посредством фагов.
История изучения
Процесс трансформации был открыт в 1928 г. английским микробиологом Ф. Гриффитом. Этот процесс Гриффит наблюдал на двух видах штаммов пневмококковых бактерий (S и R). Штамм S имел полисахаридную оболочку, поверхность клетки была гладкой, а штамм R не имел полисахаридной оболочки, и поверхность клетки была шероховатой (буква S — от англ. smooth — гладкий, R — от англ. rough — шероховатый). Полисахаридная оболочка штамма S бактерии вызвала пневмококковое заболевание и привела к гибели животных. Штамм R не был возбудителем болезни, поэтому мыши, заражённые этим штаммом, не погибли. При нагревании болезнетворных S-штаммов они погибали, и при введении клеток убитого S-штамма мышам не вызывали заболевания. При введении мышам убитых нагреванием S-штаммов бактерий вместе с неболезнетворными живыми клетками R-штамма животные погибали. В крови погибших мышей были найдены живые клетки S- штамма бактерий. Сущность этого явления показана на рис. 4. Видно, что какое-то вещество переходит из клеток S-штамма пневмококка в R-штамм и изменяет его наследственность. Под воздействием этого вещества отдельные клетки R-штамма превратились в S-штамм, то есть трансформировались. Гриффит не смог объяснить полученные в опытах результаты.
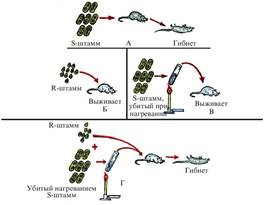
Описание к опыту Гриффита (рис. 4). А — мышь, заражённая патогенной (болезнетворной) пневмококковой бактерией (S-штамм), погибает; Б — мышь, заражённая непатогенной (R-штамм) пневмококковой бактерией, выживает; В — мышь, заражённая патогенной бактерией, убитой нагреванием, выживает; Г — при смешивании S- и R-штаммов бактерий и введении этой смеси мыши она погибает. В её крови обнаруживается живой 5-штамм. Это свидетельствует о том, что при введении в организм мыши смеси, состоящей из непатогенного R- и убитого S-штаммов, ген наследственности молекулы S-штамма, вызывающий болезнь, переносится в наследственную молекулу живого R-штамма и придаёт ей свои свойства, т. е. происходит трансформация.
Эксперимент Херши—Чейз
Продолжение 8 вопроса
Репликация – это процесс удвоения ДНК, в процессе репликации каждая цепь материнской ДНК служит матрицей длясинтезы новой дочерней цепи. Репликация происходит в S-фазу клеточного цикла. Хромосома содержит 1ну двухцепочечную молекулу ДНК.
Основы для репликации:
1. Принцип комплементарности (взаимосоответствия) – способность формировать водородные связи.
2. Матричный принцип – каждая из 2х нитей – матрица для создания комплементарной дочерней цепи.
3. Полуконсервативный механизм – при удвоении ДНК новая двухцепочечная молекула содержит 1ну исходную цепь и 1ну вновь синтезированную.
Условия репликации:
1) Матрица – 2 расплетенные нити ДНК, образующие репликационную вилку.
2) Субстраты и источники энергии для синтеза – дНТФ (дАТФ, дГТФ, дЦТФ, дТТФ)
3) Ионы Mg
4) Репликативный комплекс ферментов:
ДНК-хеликаза– расплетает двойную спираль ДНК и разрывает водородные связи.
ДНК-топоизомераза 1и2 –устраняют суперспирализацию ДНК в точке разрыва, облегчая формирование репликационной вилки.
SSB-белки –связываются с одноцепочечными нитями ДНК и препятствуют их повторному комплементарному соединению.
ДНК-полимераза – катализирует образование фосфодиэфирных связей между нуклеотидами и удлинение уже существующей цепи. 2 свободных нуклеотида она соединять не может.
Праймаза– катализирует синтез олигорибонуклеотидов – коротких молекул РНК.
ДНК-лигаза – соединяет фрагменты ДНК.
Теломераза– обеспечивает построение концов ДНК.
Этапы репликации:
Инициация – формирование репликативной вилки при участии ДНК-хеликазы и ДНК-топоизомераз, в итоге происходит разделение цепей ДНК, а приссоединенные SSB-белки препятствуют повторному соединению цепей. Инициация происходит в нескольких точках – точках инициации или ориджины репликации. Единица репликации эукариот - репликон – участок между соседними ориджинами.
Элонгация – синтез новой цепи ДНК, она начинается праймазой, которая синтезирует олигорибонуклеотиды – праймеры. Далее синтез дочерней цепи ДНК происходит за счет полимеразы от 5` к 3` концу. Новые цепи синтезируются неодинаково. На 1ой цепи ДНК 5`- 3`цепь растет непрерывно, быстро и называется лидирующей. На другой нити ДНК новая цепь синтезируется в виде фрагментов Оказаки – эта цепь называется отстающей.
Терминация – удаление праймеров и завершение формирования отстающей цепи ДНК. Праймеры удаляются эндонуклеазой, а на их месте ДНК-полимераза достраивает ДНК. Сшивание фрагментов происходит за счет ДНК-лигазы. В итоге на каждой матричной ДНК формируется 2 дочернии копии ДНК.
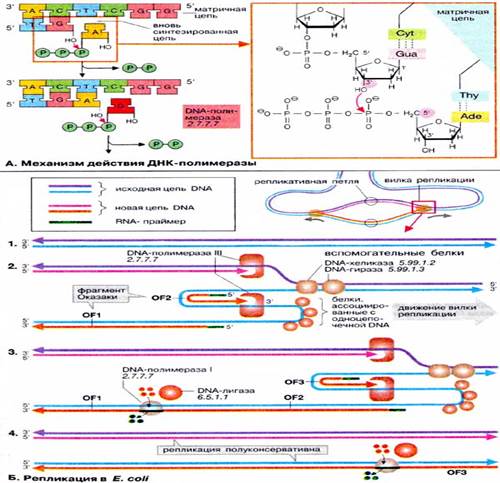
Б. Репликация в Е. coli
В настоящее время процесс репликации у прокариот достаточно изучен, в то время как многие аспекты эукариотической репликации остаются неясными. Однако с большой долей вероятности можно утверждать, что в большинстве клеток этот процесс протекает в основном одинаково. На схеме показана простейшая схема репликации у бактерии Escherichia coli. В бактериях репликация начинается со специфической точки в кольцевой ДНК (область начала репликации) и продолжается в обоих направлениях. В результате образуются две репликативные вилки, которые продвигаются в противоположных направлениях, т. е. обе цепи реплицируются одновременно. На схеме исходная ДНК (1) окрашена в голубой и фиолетовый цвета, а вновь синтезирующаяся — в розовый и оранжевый. В функционировании каждой вилки принимают участие множество различных белков, из которых здесь указаны наиболее важные.
Каждая репликативная вилка (2) включает по крайней мере две молекулы ДНК-полимеразы III, ассоциированные с несколькими вспомогательными белками. К последним относятся ДНК-топоизомеразы (гиразы), которые раскручивают плотно свернутую двойную спираль ДНК, и хеликазы, которые расплетают двухтяжевую ДНК на две цепи. Поскольку матричная цепь всегда читается в направлении 3'→5' (см. выше), только одна из цепей может считываться непрерывно (розовая/фиолетовая; 2). Другая цепь (голубого цвета) считывается в направлении, противоположном движению репликативной вилки. В результате на матрице вначале синтезируются короткие фрагменты новой цепи ДНК (зеленый/оранжевый), так называемые фрагменты Оказаки (OF), названные так по имени их первооткрывателя. Каждый фрагмент начинается с короткой РНК-затравки (праймера, зеленого цвета), необходимой для функционирования ДНК-полимеразы. Праймер синтезируется специальной РНК-полимеразой («праймаза», на схеме не показана), ДНК-полимераза III достраивает этот праймер до фрагмента ДНК длиной 1000-2000 дезоксинуклеотидных звеньев (оранжевого цвета). Синтез этого фрагмента далее прерывается, и новый синтез начинается со следующего РНК-праймера. Индивидуальные фрагменты Оказаки первоначально не связаны друг с другом и все еще имеют РНК на 5'-концах (3). На некотором расстоянии от репликативной вилки ДНК-полимераза I начинает замещать РНК-праймер последовательностью ДНК. В завершение остающиеся одноцепочечные разрывы репарируются ДНК-лигазой. В образованной таким образом двойной спирали ДНК только одна из цепей синтезирована заново. Поэтому говорят, что репликация ДНК происходит по полуконсервативному механизму.
История изучения
Попытки понять, каким образом последовательность ДНК кодирует аминокислотную последовательность белков, начали предприниматься почти сразу же после того, как в 1953 году была установлена структура ДНК (двойная спираль[en]). Георгий Гамов предположил, что кодоны должны состоять из трёх нуклеотидов, чтобы кодонов хватило для всех 20 аминокислот (всего же возможно 64 различных кодона из трёх нуклеотидов: на каждую из трёх позиций можно поставить один из четырёх нуклеотидов),
В 1961 году триплетность генетического кода удалось подтвердить экспериментально. В том же году Маршалл Ниренберг и его коллега Heinrich Matthaei использовали бесклеточную систему для трансляции in vitro. В качестве матрицы был взят олигонуклеотид, состоящий из остатков урацила (UUUU...). Пептид, синтезированный с него, содержал только аминокислоту фенилаланин. Так впервые было установлено значение кодона: кодон UUU кодирует фенилаланин. Дальнейшие правила соответствия между кодонами и аминокислотами были установлены в лаборатории Северо Очоа. Было показано, что полиадениновая РНК (ААА...) транслируется в полилизиновый пептид, а на матрице полицитозиновой РНК (ССС...) синтезируется пептид, состоящий только из остатков пролина. Значение остальных кодонов было установлено при помощи разнообразных сополимеров в ходе экспериментов, проведённых в лаборатории Хара Гобинда Кораны. Вскоре после этого Роберт Холли установил структуру молекулы тРНК, которая служит посредником при трансляции. В 1968 году Ниренберг, Корана и Холли были удостоены Нобелевской премии по физиологии и медицине.
Ответ на 4 вопрос
Трансформацией называется перенос в определённых условиях любой части наследственной молекулы одного организма в наследственную молекулу другого организма. Другими словами — это переход отрезка ДНК одной клетки в геном другой клетки в функционально активном состоянии.
Процесс трансформации представляет собой естественный процесс, который приводит к изменению наследственности клеток.
Изучение процесса трансформации послужило толчком для открытия трансдукции — процесса перенос генов из одной бактериальной клетки в другую посредством фагов.
История изучения
Процесс трансформации был открыт в 1928 г. английским микробиологом Ф. Гриффитом. Этот процесс Гриффит наблюдал на двух видах штаммов пневмококковых бактерий (S и R). Штамм S имел полисахаридную оболочку, поверхность клетки была гладкой, а штамм R не имел полисахаридной оболочки, и поверхность клетки была шероховатой (буква S — от англ. smooth — гладкий, R — от англ. rough — шероховатый). Полисахаридная оболочка штамма S бактерии вызвала пневмококковое заболевание и привела к гибели животных. Штамм R не был возбудителем болезни, поэтому мыши, заражённые этим штаммом, не погибли. При нагревании болезнетворных S-штаммов они погибали, и при введении клеток убитого S-штамма мышам не вызывали заболевания. При введении мышам убитых нагреванием S-штаммов бактерий вместе с неболезнетворными живыми клетками R-штамма животные погибали. В крови погибших мышей были найдены живые клетки S- штамма бактерий. Сущность этого явления показана на рис. 4. Видно, что какое-то вещество переходит из клеток S-штамма пневмококка в R-штамм и изменяет его наследственность. Под воздействием этого вещества отдельные клетки R-штамма превратились в S-штамм, то есть трансформировались. Гриффит не смог объяснить полученные в опытах результаты.
Описание к опыту Гриффита (рис. 4). А — мышь, заражённая патогенной (болезнетворной) пневмококковой бактерией (S-штамм), погибает; Б — мышь, заражённая непатогенной (R-штамм) пневмококковой бактерией, выживает; В — мышь, заражённая патогенной бактерией, убитой нагреванием, выживает; Г — при смешивании S- и R-штаммов бактерий и введении этой смеси мыши она погибает. В её крови обнаруживается живой 5-штамм. Это свидетельствует о том, что при введении в организм мыши смеси, состоящей из непатогенного R- и убитого S-штаммов, ген наследственности молекулы S-штамма, вызывающий болезнь, переносится в наследственную молекулу живого R-штамма и придаёт ей свои свойства, т. е. происходит трансформация.
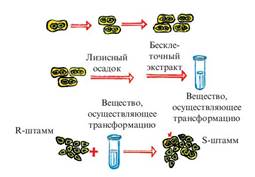
Эксперимент Эвери, Маклеода и Маккарти
Данное явление было объяснено в 1944 г. О. Эвери с сотрудниками Маклеодом и Маккарти. Для этого они расщепили пневмококковую клетку S-штамма на отдельные фракции, проверили болезнетворную активность каждой фракции. Было установлено, что только при смешивании выделенной из S-штамма молекулы ДНК с R-штаммом непатогенный R-штамм трансформируется в вызывающий болезнь S-штамм (рис. 5). Следовательно, было доказано, что трансформация R-штамма в S-штамм зависит от молекулы ДНК.
Эксперимент Херши—Чейз
Дата: 2019-02-02, просмотров: 560.