| Представители животного мира | Разрушение передних (двигательных) отделов коры | Разрушение задних (сенсорных) отделов коры |
| Птица | Продолжает летать; изменений в движениях не заметно | Четко ориентируется; выбирает площадку, на которую садится |
| Собака | Движения конечностей, противоположных очагу разрушения, нарушаются; паралич противоположных конечностей, который, однако, частично претерпевает обратное развитие | Частично страдают реакции на экстероцептивные стимулы |
| Обезьяна | Стоите помощью | Значительное нарушение процессов восприятия (частично восстанавливающихся) |
| Человек | Полный и стойкий паралич конечностей, противоположных очагу разрушения | Дифференцированное и необратимое нарушение отдельных форм чувствительности |
Другая причина противоречивости результатов стала ясна из сравнительного анализа эффектов разрушения коры головного мозга. Оказалось, что разрушение коры головного мозга у птиц (у которых кора едва намечена) приводит к относительно незначительным результатам; у мыши подобное разрушение вызывает также небольшие изменения поведения; у собаки объем этих изменений возрастает, и они делаются более стойкими; у обезьяны дифференцированность и стойкость нарушений поведения, возникших в результате разрушения ограниченных участков мозга, становится несравненно более отчетливой; у человека локальные разрушения мозга вызывают стойкие нарушения различных психических процессов.
Таким образом, на последовательных ступенях эволюции поведение животного в разной степени зависит от высших отделов мозга (в частности от его коры): чем выше на эволюционной лестнице стоит животное, тем в большей степени его поведение регулируется корой и тем больше возрастает дифференцированный характер этой регуляции. Этот закон прогрессивной кортикализации функций — один из основных законов мозговой организации поведения — является ключом к пониманию причины тех противоречий, которые наблюдали отдельные авторы, изучавшие мозговую организацию поведения животных.
Роль коры головного мозга животных в организации специальных видов поведения была хорошо изучена известным американским психологом К. С.Лешли и великим русским физиологом И. П. Павловым. Следующий пример дает возможность проиллюстрировать те факты, с которыми столкнулся первый из этих исследователей, и показать трудности, связанные с решением вопроса о локализации сложных психических функций в коре головного мозга животных.
Изучая мозговую организацию зрения животных, Лешли (1930— 1942) обнаружил, что разрушение затылочной области коры приводит к тому, что обезьяна, продолжая без труда различать звуки и прикосновения, теряет возможность различать зрительно предъявляемые фигуры. Эти факты, многократно подтвержденные другими авторами (Клювер, 1927, 1937, 1941), с достаточной убедительностью показали, что задневисочные и затылочные отделы мозга этих животных имеют прямое отношение к организации зрительного восприятия и что разрушение этих областей мозговой коры неизбежно приводит к нарушению сложных форм анализа зрительных раздражений. Несмотря на всю убедительность этих данных, Лешли столкнулся с парадоксальным фактом. Оказалось, что обезьяна, лишенная зрительной (затылочной) коры, продолжала с успехом отличать треугольник от круга, успешно выполняя тем самым функции, которые, казалось бы, должны были распадаться при разрушении этого высшего аппарата зрительного восприятия.
Дальнейший анализ позволил, однако, убедиться в неправильности такого вывода: оказалось, что обезьяна, лишенная зрительной коры, различает не форму, а общую массу освещенности, различную у обеих фигур; поэтому стоило только уравнять массу света, которая отражалась обеими фигурами (уравняв площадь этих фигур), чтобы их различение стало недоступным животным с разрушенной зрительной корой.
Эти факты еще раз показывают — но на этот раз в результате применения иных методов, — что в коре головного мозга животного существуют зоны, играющие решающую роль в организации сложных форм зрительного восприятия, в то время как более простые функции (различение массы света) осуществляются элементарными механизмами подкорковых зрительных ядер.
63
Позднейшие исследования Л.Вейзкранца (1964, 1968) позволили уточнить роль сенсорных отделов мозговой коры животных в организации зрительной, слуховой и проприоцептивной информации.
Внутренние физиологические механизмы функций коры головного мозга животных по анализу и переработке сенсорной информации были с полной отчетливостью установлены работами школы И.П.Павлова, которые хронологически предшествовали всем только что упомянутым исследованиям.
Еще в первом десятилетии нашего века (XX в. — примеч. ред.) сотрудники И. П. Павлова установили фундаментальный факт: разрушение определенной области коры головного мозга собаки может привести к нарушению аналитико-синтетической работы соответствующего анализатора, не затрагивая, однако, наиболее простых форм сенсорных процессов. Так, было установлено, что собаки с удаленной височной областью коры могут реагировать на звуки, но оказываются не в состоянии выработать дифференцированные системы условных рефлексов на комплексные звуковые раздражители (Б.П.Бабкин, 1910; А.Н.Кудрина, 1910; И.Н.Кржыжановский, 1909; М. И. Эльяссон, 1908; и др.); те же закономерности были выявлены в отношении зрительного и кожного анализаторов.
Все эти исследования дали возможность убедиться в том, что кора головного мозга является аппаратом, обеспечивающим не столько элементарные сенсорные функции, сколько сложный процесс анализа и синтеза поступающей информации, и сделать тем самым шаг вперед в понимании основных физиологических законов этой сложнейшей деятельности.
Клинический материал о нарушениях психической деятельности мозга человека настолько богат и занимает такое важное место в построении нейропсихологии, что мы не будем рассматривать его здесь и посвятим его анализу специальную главу.
Хирургическое разрушение отдельных участков мозга — метод, которым в течение ряда поколений пользовались физиологи для изучения функций отдельных систем мозга, является далеко не идеальным методом исследования. Каждое хирургическое вмешательство вызывает кровоизлияние (с последующим образованием рубцов) и, таким образом, — обширные изменения мозговой ткани, что существенно затрудняет анализ роли разрушенного участка коры в построении соответствующей функции.
Вот почему в последнее время стали успешно применяться другие — бескровные — методы выключения определенных участков мозга. К числу их относится местное охлаждение отдельных участков мозга, смазывание отдельных зон мозга алюминиевой пастой (Прибрам, 1960, 1969) и, наконец, воздействие на участки мозга постоянным током (Спинелли и Прибрам, 1967), нарушающим их нормальное функционирование.
64
Преимущество этих методов перед методами разрушения состоит не только в том, что они не дают побочных, перифокальных явлений, но и в том, что такое выключение носит временный характер и что наряду с последствиями выключения может быть прослежен и процесс обратного включения соответствующих участков коры в действующие мозговые системы.
Данные, полученные благодаря применению этих методов, подтверждают и существенно обогащают те основные положения, на которых мы останавливались ранее.
Совокупность фактов, полученных в сравнительно-анатомических исследованиях, при изучении особенностей поведения, а также в физиологических работах, основанных на методе раздражения или методе выключения (разрушения) отдельных участков мозга, позволяет нам прийти к ряду решающих и в целом совершенно однозначных выводов.
Все эти источники показали, что по мере эволюции животного мира поведение все больше зависит от высших этажей мозга (его коры) или, другими словами, что на высших ступенях эволюции процесс прогрессивной кортикализации становится все более отчетливым. Кроме того, по мере эволюции функциональная организация наиболее высоких аппаратов мозговой коры становится все более дифференцированной и каждая система большого мозга приобретает отчетливую иерархическую организацию, едва намеченную у низших позвоночных, но становящуюся ведущей характеристикой мозга у приматов и особенно у человека.
Все использованные наукой источники подтверждают, что каждая из действующих систем головного мозга (зрительная, слуховая, общечувствительная и двигательная) имеет вергикильную организацию, начинаясь периферическими рецепторами, переходящими в проводящие пути, включающие наиболее простые — интегрирующие — аппараты верхнего ствола и межуточного мозга, и кончаясь высокодифференцированными аппаратами мозговой коры.
Вместе с тем источники показали, что каждая из этих систем имеет иерархическое строение; система состоит из группы надстроенных друг над другом корковых зон. В основе каждой системы лежат первичные (или проекционные) зоны коры, куда приходят импульсы, полученные посредством периферических рецепторов (органов чувств), и откуда на периферию направляются двигательные импульсы. Эти аппараты коры дробят воспринимаемую информацию на миллионы составляющих ее признаков и тем самым делают доходящие до коры возбуждения доступными кодированию и управлению.
65
Над первичными зонами надстроены вторичные зоны коры, способные благодаря преобладанию в них верхних (ассоциативных) слоев нейронов к анализу и синтезу поступающей информации, к переработке (кодированию) и хранению материала чувственного опыта и к подготовке сложных двигательных программ. Вторичные зоны коры, связанные с периферией посредством ассоциативных ядер зрительного бугра, составляют аппарат обеспечения синтетических форм работы отдельных анализаторов и занимают в коре головного мозга человека важнейшее место.
Наконец, над всем этим комплексом корковых аппаратов специализированного (модально-специфического) синтеза надстраиваются третичные зоны коры, которые выделяются в процессе эволюции позднее других и приобретают решающее значение только у человека.
Эти зоны, обладающие особенно тонким и сложным строением и располагающие мощным аппаратом ассоциативных нейронов верхних слоев коры, находятся либо на стыке вторичных отделов зрительного, слухового и общечувствительного анализаторов (вследствие чего они получили название «зон перекрытия»), либо же в передних (префронтальных) отделах большого мозга, сохраняя связь со всеми остальными отделами коры.
Изучение этих структур показывает, что они играют особенно важную роль в функциональной организации мозга, обеспечивая совместную работу отдельных анализаторов и тем самым образуя основу для получения целостной картины мира. Они являются также мозговым аппаратом, ответственным за формирование планов и программ поведения, регуляцию и контроль человеческой деятельности.
Роль третичных аппаратов мозга в человеческом поведении настолько велика, что описание их функций будет предметом рассмотрения ряда разделов этой книги.
Глава II
ЛОКАЛЬНЫЕ ПОРАЖЕНИЯ МОЗГА И ОСНОВНЫЕ ПРИНЦИПЫ ЛОКАЛИЗАЦИИ ФУНКЦИЙ
Предыдущая глава была посвящена основным источникам наших знаний о функциональной организации мозга, в ней, в частности, мы познакомились с теми данными, которые наука получила в опытах с выключением (разрушением) отдельных зон коры головного мозга животных. Однако эти данные имеют несравнимо меньшее значение для развития современных представлений о мозге как органе психики, чем результаты наблюдений над изменениями психических процессов человека при ограниченных (локальных) поражениях мозга.
Вот почему нейропсихологическое изучение больных с локальными поражениями мозга с полным основанием может считаться основным источником современных представлений о функциональной организации мозга как органа психической жизни и почему мы считаем необходимым рассмотреть получаемые этим путем факты в особой главе.
РАННИЕ РЕШЕНИЯ
Попытки рассматривать сложные психические процессы как функцию ограниченных участков мозга относятся к еще очень давнему времени. В средние века философы и натуралисты считали возможным локализовать сложные «психические способности» в трех мозговых желудочках (рис. 31). В самом начале XIX в. известный анатом Ф. Галль, впервые описавший различия между серым и белым веществом головного мозга, высказал мысль, что сложные «способности» человека интимно связаны с отдельными, строго ограниченными, участками мозга, которые, разрастаясь, образуют соответствующие выпуклости на черепе, позволяющие определять индивидуальные различия в способностях человека.
Рис. 31. Карты «трех желудочков
мозга»: 1 — воображение;
2 — мышление; 3 — память
67
Френологические карты Ф. Галля (рис. 32), представлявшие собой, по сути дела, ни на чем не основанную попытку спроецировать на мозг современную ему «психологию способностей», очень быстро были забыты. На смену им пришли попытки выделить функциональные зоны мозговой коры на основе наблюдений над изменениями в поведении человека в результате ограниченных поражений мозга.
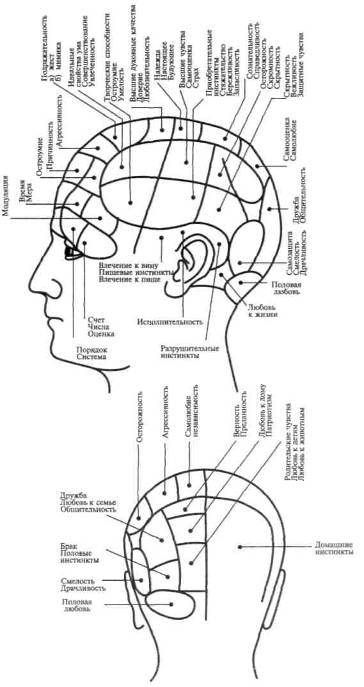
Рис. 32. Френологические карты Ф.Галля
Клинические наблюдения больных с локальными поражениями мозга начались давно; довольно рано было установлено, что поражение двигательной зоны коры ведет к параличу противоположных конечностей, поражение заднецентральной области коры — к выпадению чувствительности на противоположной стороне тела, а поражение затылочной области мозга — к центральной слепоте.
Однако подлинное рождение учения о нарушении психических процессов с полным основанием можно отнести к 1861 году, когда молодой французский анатом П. Брока описал мозг больного, который в течение многих лет страдал грубым нарушением моторной (экспрессивной) речи; Брока установил, что в мозгу этого больного была разрушена задняя треть нижней лобной извилины. Через несколько лет дополнительные наблюдения позволили П. Брока показать, что моторная речь связана с ограниченной областью головного мозга, а именно — с задней третью нижней лобной извилины левого полушария.
Так как у всех наблюдавшихся им больных были сохранны как понимание речи, так и движения губ и языка, П. Брока предположил, что задняя треть нижней лобной извилины левого полушария является «центром моторных образов слов» и что поражение этой области приводит к своеобразному выпадению экспрессивной речи, которую он сначала назвал «афемией» и лишь потом заменил это название на существующий ныне термин афазия.
Открытие Брока имело двойной смысл. С одной стороны, впервые сложная психическая функция была локализована в определенном участке мозговой коры, причем эта локализация в отличие от фантастических попыток Ф. Галля, на поколение раньше Брока предложившего свою «френологию», носила клинически обоснованный характер.
С другой стороны, это открытие впервые показало коренное различие между функциями левого и правого полушарий мозга, выделив левое полушарие (у правшей) как ведущее, связанное со сложнейшими речевыми функциями.
68
Потребовалось лишь одно десятилетие, чтобы плодотворность открытия Брока стала очевидной: в 1873 г. немецкий психиатр К. Вернике описал случай, когда поражение другого участка (задней трети верхней височной извилины левого полушария) вызывало столь же четкую, но на этот раз обратную картину — нарушение понимания слышимой речи при относительной сохранности экспрессивной (моторной) речи. Развивая идеи П. Брока, Вернике предположил, что задняя треть первой височной извилины левого полушария является «центром сенсорных образов слова» или, как он тогда выражался, центром «понятия слова» («Wortbegriff»).
Открытие того факта, что сложнейшие формы психической деятельности могут рассматриваться как функции ограниченных участков мозга, иначе говоря, что они могут быть локализованы в ограниченных областях мозговой коры так же, как и элементарные функции (движения, чувствительность), вызвало небывалое оживление в неврологической науке, и невропатологи стали с энтузиазмом собирать факты, подтверждающие, что и другие сложные психические процессы являются функцией отдельных участков мозговой коры.
В результате такого бурного интереса к прямой локализации функций в ограниченных зонах мозговой коры в течение очень короткого срока — «блистательных 70-х годов» — в коре головного мозга были найдены «центр понятий» (в нижнетеменной области левого полушария), «центр письма» (в задних отделах средней лобной извилины этого же полушария), «центры счета», «центры чтения», «центры ориентировки в пространстве» и т.д. Позднее были описаны аппараты связей между ними, и к 80-м годам XIX в. неврологи и психиатры стали составлять «функциональные карты» мозговой коры, которые, как им казалось, окончательно разрешали вопрос о функциональном строении мозга как органа психической деятельности. Дальнейшее накопление материала благодаря наблюдениям над последствиями очаговых (локальных) поражений различных участков мозга вследствие мозговых ранений или местных кровоизлияний стимулировало эти попытки, и тенденция локализовывать сложнейшие психологические процессы в ограниченных участках мозга сохранялась в течение более чем полувека.
Данные «узких локализационистов», наблюдавших последствия ограниченных поражений мозговой коры (выпадение узнавания чисел, нарушение понимания слов или фраз, нарушение узнавания предметов, нарушение побуждений, изменения личности и т.д.), не опосредствованные тщательным психологическим анализом наблюдаемых симптомов, позволили им создать целый ряд новых гипотетических карт локализации функций в коре голов-
70
ного мозга. Наиболее подробные из таких карт были предложены немецким психиатром Клейстом (1934), который на основании обработки огромного материала наблюдений над огнестрельными ранениями мозга в течение Первой мировой войны разместил в различных участках мозговой коры такие «функции», как «схема тела», «понимание фраз», «конструктивные действия», «настроение» и т.д. (рис. 33). Принципиально такие карты немногим отличались от френологических карт Ф. Галля.
Рис. 33. Локализационная карта Клейста: вверху — наружная поверхность; внизу — внутренняя поверхность мозга
Попытки непосредственной локализации сложных психических функций в ограниченных участках мозга были настолько стойкими, что еще в 1946 г. известный американский невролог Нильсен считал возможным описывать ограниченные участки коры, которые, по его мнению, были «центрами восприятия живых объектов», отличая их от других участков, где локализовалось восприятие «неживых объектов».
2 КРИЗИС
Было бы, однако, неверным думать, что стремлениями к непосредственной локализации сложных психологических процессов в ограниченных участках мозга, или «узким локализационизмом», как его принято называть, исчерпывалось развитие неврологической мысли и что это направление не встречало противодействия со стороны достаточно влиятельных неврологов.
Уже в «блистательные 70-е годы» Брока и следовавшие за ним ученые встретили мощного оппонента в лице известного английского невролога Хьюлингса Джексона, высказавшего предположение, что к мозговой организации сложных форм психических процессов следует подходить скорее с позиций уровня их построения, чем с позиций их локализации в ограниченных участках мозга.
Положения Джексона оказались слишком сложными для его времени — лишь много лет спустя они были подхвачены и развиты выдающимися неврологами первой половины двадцатого века — Монаковым (1914), Хэдом (1926) и Гольдштейном (1927, 1934, 1948). Не отрицая того очевидного факта, что элементарные физиологические функции (такие, как кожная чувствительность, зрение, слух, движение) обеспечиваются четко очерченными участками коры головного мозга, эти исследователи выразили справедливое сомнение в том, что принцип узкой локализации приложим к мозговым механизмам сложных форм психической деятельности.
72
Указывая на сложный характер психической деятельности человека и пытаясь найти его специфические черты в осмысленном характере поведения (Монаков) или абстрактной установке и категориальном поведении (Гольдштейн), эти авторы не принимали положения о том, что соответствующие функции привязаны к ограниченным участкам мозга так же, как элементарные отправления мозговой ткани, и высказывали мысль, что сложные явления семантики или категориального поведения являются, скорее, результатом деятельности всего мозга, чем продуктом работы ограниченных участков мозговой коры. Сомнения в возможности узкой локализации сложных психических процессов приводили этих авторов либо к отрыву психических процессов от мозгового субстрата и к признанию их особой духовной природы, а именно такую позицию заняли к концу своей жизни такие выдающиеся исследователи, как Монаков (1928) и Шеррингтон (1934, 1942), либо же к попытке показать, что категориальное поведение является наиболее высоким уровнем мозговой деятельности, зависящим в большей степени от массы вовлеченного в работу мозга, чем от участия тех или иных определенных зон мозговой коры (Гольд-штейн, 1934, 1948). Таким образом, справедливые сомнения в правомерности механистического подхода «узких локализационистов» приводили либо к оживлению дуалистического признания духовной природы психических процессов, либо к оживлению идей о мозге как недифференцированном целом и решающей роли его массы в осуществлении психической деятельности, неоднократно всплывавших на протяжении всей истории изучения мозга (Флуранс, 1824; Гольц, 1876-1884; Лешли, 1929).
Легко увидеть, что если механистические представления о прямой локализации психических процессов в ограниченных участках мозга приводили исследования мозговых основ психической деятельности к тупику, то и «целостные» (или, как их иногда принято называть, «ноэтические») представления о психических процессах не создавали нужной основы для дальнейших научных исследований, сохраняя отжившие представления об обособленности духовной жизни человека и принципиальной невозможности искать ее материальную основу, равно как и столь же отжившие представления о мозге как о примитивной, недифференцированно-нервной массе.
Совершенно естественно, что такой кризис создавал необходимость поиска новых путей, которые позволили бы выявить подлинные мозговые механизмы психической деятельности человека, опираясь на те же научные принципы исследования, которые оправдали себя при изучении элементарных форм физиологических процессов, но которые были бы адекватными объекту изучения — сознательной деятельности человека, социально-исторической по своему происхождению и сложной, опосредствованной по своему строению.
Такая задача требовала коренной перестройки понимания функций, с одной стороны, и основных принципов их локализации — с другой.
73
ПЕРЕСМОТР ОСНОВНЫХ ПОНЯТИЙ
Для того чтобы подойти к вопросу о мозговой локализации сложных форм психической деятельности человека, нам нужно прежде всего остановиться на том пересмотре основных понятий, без которого правильное решение этого вопроса останется невозможным. Мы имеем в виду прежде всего пересмотр понятия «функция», затем — понятия «локализация» и, наконец, переоценку того, что называлось «симптомом», или «выпадением» функции при локальном поражении мозга.
Пересмотр понятия «функция»
Исследователи, пытавшиеся рассмотреть вопрос о локализации элементарных функций в коре головного мозга, пользуясь как методом раздражения, так и методом выключения ограниченных участков мозга, понимали «функцию» как отправление той или иной ткани.
Такое понимание, несомненно, правомерно. Совершенно естественно считать, что выделение желчи есть функция печени, а выделение инсулина — функция поджелудочной железы. Столь же правомерно рассматривать восприятие света как функцию светочувствительных элементов сетчатки глаза и связанных с нею высокоспециализированных нейронов зрительной коры, а генерацию двигательных импульсов — как функцию гигантских пирамидных клеток Беца.
Однако такое определение не исчерпывает всех аспектов понятия «функция».
Когда мы говорим о функции пищеварения или функции дыхания, понимание ее как отправление определенной ткани становится явно недостаточным.
Для осуществления акта пищеварения требуется доведение пищи до желудка, переработка пищи под влиянием желудочного сока, участие в этой переработке секретов печени, поджелудочной железы, сокращение стенок желудка и кишечника, проталкивание усваиваемого вещества по пищевому тракту и, наконец, всасывание расщепленных элементов пищи стенками тонкого кишечника.
Точно так же обстоит дело с функцией дыхания. Конечной задачей дыхания является доведение кислорода до легочных альвеол и его диффузия через стенки альвеол в кровь. Однако для реализации этой конечной цели необходимо участие сложного мышечного аппарата, в состав которого входят мышцы диафрагмы и межреберные мышцы, позволяющие расширять и сужать объем грудной клетки и управляемые сложнейшей системой нервных приборов ствола мозга и вышележащих образований.
74
Понятно, что весь этот процесс представляет собой не просто функцию, а целую функциональную систему, включающую многие звенья, расположенные на различных этажах секреторного, двигательного и нервного аппаратов.
Такая «функциональная система» (термин, введенный П.К.Анохиным, 1935, 1940, 1949, 1963, 1968; и др.) отличается не только сложностью строения, но и подвижностью входящих в ее состав частей.
Легко видеть, что исходная задача (восстановление гомеоста-за) и конечный результат (доведение питательных веществ до стенок кишечника или кислорода до альвеол) остаются во всех случаях одинаковыми (или, как иногда говорят, инвариантными). Однако способ выполнения этой задачи может сильно варьировать. Так, если основная группа работающих при дыхании мышц диафрагмы перестает действовать, в работу включаются межреберные мышцы, а если и они почему-либо страдают, включаются мышцы гортани и воздух как бы заглатывается животным. Наличие постоянной (инвариантной) задачи, осуществляемой с помощью меняющихся (вариативных) средств, позволяющих доводить процесс до постоянного (инвариантного) результата, является одной из основных особенностей работы каждой функциональной системы.
Другой особенностью является сложный состав функциональной системы, всегда включающей целый набор афферентных (настраивающих) и эфферентных (осуществляющих) компонентов.
Такое представление о функции как о целой функциональной системе резко отлично от представления о ней как об отправлении определенной ткани.
Если уж наиболее сложные вегетативные и соматические процессы построены по типу таких функциональных систем, то с еще большим основанием это понятие можно отнести к сложным функциям поведения.
Проиллюстрируем это на примере функции движения (локомо-ции), детальная структура которой была разработана замечательным отечественным психофизиологом Н.А. Бернштейном (1935, 1947, 1957, 1966).
Движение человека, имеющего намерение переместиться в пространстве, попасть в определенную точку или выполнить некоторое действие, никогда не может осуществиться одними лишь эфферентными, двигательными импульсами. Уже то, что опорный двигательный аппарат с подвижными суставами имеет, как правило, огромное число степеней свободы, умножающееся еще более от того, что в движении участвуют группы сочленений, делает движение принципиально неуправляемым одними только эфферентными импульсами. Для того чтобы начавшееся движение осуществилось, необходима постоянная коррекция его афферентными импульсами, которые сигнализируют о положении движущейся конечности в пространстве и об изменении вязкости мышц.
75
Только такое сложное строение двигательного процесса может обеспечить выполнение постоянной (инвариантной) двигательной задачи меняющимися (вариативными) средствами, т.е. достижение с помощью этих динамически меняющихся средств постоянного (инвариантного) полезного результата.
В известных опытах Хантера мышь в лабиринте достигала цели путем побежки, а когда одно из звеньев лабиринта заменялось водным бассейном, — посредством плавательных движений. В экспериментах Лешли крыса, натренированная проходить определенный путь, коренным образом меняла состав движений, когда у нее удалялся мозжечок или когда ее спинной мозг рассекался двумя половинными разрезами так, что ни одно волокно не достигало периферии, — в этих случаях, потеряв способность воспроизводить хорошо заученные движения, она достигала цели (нужного результата), двигаясь «кубарем».
Сменный характер движений, необходимых для осуществления цели, выступает также и при тщательном анализе любого двигательного акта человека: попадание в цель, например, может осуществляться различным набором движений в зависимости от исходного положения тела, другой пример — процесс письма, который можно осуществлять карандашом или ручкой, правой рукой или левой и даже ногой, не изменяя при этом ни содержания написанного, ни даже характерного для пишущего почерка (Н.А. Бернштейн, 1947).
Такое системное строение, характеризующее относительно простые поведенческие акты, в неизмеримо большей степени характерно для более сложных форм психической деятельности.
Совершенно естественно, что такие психические процессы, как восприятие и запоминание, гнозис и праксис, речь и мышление, письмо, чтение и счет, не являются изолированными и неразложимыми «способностями» и не могут рассматриваться как непосредственные функции ограниченных клеточных групп, локализованные в определенных участках мозга.
Психические процессы, как известно, формировались в течение длительного исторического развития. Будучи социальными по своему происхождению и сложными, опосредствованными по строению, они опираются на сложную систему способов и средств. Эти положения, подробно разработанные в трудах выдающегося представителя отечественной психологической науки Л.С.Выготского (1956, 1960), а также его учеников — А.Н.Леонтьева (1959), А.В.Запорожца (1960), П.Я.Гальперина (1959), Д. Б. Эльконина (1960) и др., заставляют относиться к основным формам сознательной деятельности как к сложнейшим функциональным системам и, как следствие, коренным образом пересмотреть проблему локализации их в коре головного мозга.
76
Пересмотр понятия «локализация»
Сказанное о строении функциональных систем вообще и высших психологических функций в частности заставляет нас коренным образом пересмотреть классические представления о локализации психических функций в коре головного мозга человека.
Если элементарные отправления той или иной ткани по определению четко локализованы в тех или иных клеточных группах, то о локализации сложных функциональных систем в ограниченных участках мозга или мозговой коры, конечно, не может быть и речи.
Даже такая функциональная система, как дыхание (о которой мы уже говорили), включает в свой состав настолько много элементов, что уместно вспомнить здесь слова И.П.Павлова, сказанные им при обсуждении вопроса о «дыхательном центре»: «Если с самого начала думали, что это — точка с булавочную головку в продолговатом мозгу...», то «теперь он чрезвычайно расползся, поднялся в головной мозг и спустился в спинной, и сейчас его границы точно никто не укажет...» (Поли. собр. соч. — Т. III. — С. 127). Совершенно естественно, что с локализацией высших форм психической деятельности дело обстоит еще сложнее. Высшие формы психических процессов имеют особенно сложное строение; они складываются в процессе онтогенеза, представляя собой сначала развернутые формы предметной деятельности, которые постепенно «свертываются» и приобретают характер внутренних, умственных, действий (Л.С.Выготский, 1956, 1960; П.Я.Гальперин, 1959); как правило, они опираются на ряд внешних вспомогательных средств (язык, разрядная система счисления), сформировавшихся в процессе общественной истории, опосредствуются ими и без их участия не могут быть поняты (Л.С.Выготский, 1956, 1960), они всегда связаны с отражением внешнего мира в активной деятельности и при отвлечении от этого факта теряют всякое содержание.
Вот почему высшие психические функции как сложные функциональные системы не могут быть локализованы в узких зонах мозговой коры или в изолированных клеточных группах, а должны охватывать сложные системы совместно работающих зон, каждая из которых вносит свой вклад в осуществление сложных психических процессов и которые могут располагаться в совершенно различных, иногда далеко отстоящих друг от друга участках мозга.
Едва ли не наиболее существенным в таких системных представлениях о локализации психических процессов в коре головного мозга являются два аспекта, резко отличающие работу человеческого мозга от более элементарных форм работы мозга животного.
77
Высшие формы сознательной деятельности человека всегда опираются на внешние средства (примером могут служить узелок на платке, который мы завязываем, чтобы запомнить нужное содержание, сочетания букв, которые мы записываем, для того чтобы не забыть какую-нибудь мысль, таблица умножения, которой мы пользуемся для выполнения счетных операций, и т.п.). Эти исторически сформированные средства оказываются существенными факторами установления функциональной связи между отдельными участками мозга — с их помощью те участки мозга, которые раньше работали самостоятельно, становятся звеньями единой функциональной системы. Образно выражаясь, можно сказать, что исторически сформировавшиеся средства организации поведения человека завязывают новые «узлы» в его мозговой деятельности, и именно наличие таких функциональных «узлов», или, как некоторые называют их, «новых функциональных органов» (А. Н.Леонтьев, 1959), является важнейшей чертой, отличающей функциональную организацию мозга человека от мозга животного. Именно эту сторону построения функциональных систем человеческого мозга Л.С.Выготский (1960) называл принципом «экстракортикальной» организации сложных психических функций, имея в виду под этим не совсем обычным термином то обстоятельство, что формирование высших видов сознательной деятельности человека всегда осуществляется с опорой на ряд внешних вспомогательных орудий или средств.
Другой отличительной чертой высших психических процессов человека является тот факт, что локализация их в мозговой коре не является устойчивой, постоянной, она меняется как в процессе развития ребенка, так и на последовательных этапах упражнения. Следует пояснить это положение. Известно, что каждая сложная сознательная деятельность сначала носит развернутый характер и опирается на ряд внешних опорных средств и только затем постепенно сокращается и превращается в автоматизированный двигательный навык. Так, если на первых этапах письмо опирается на припоминание графического образа каждой буквы и осуществляется цепью изолированных двигательных импульсов, каждый из которых обеспечивает выполнение лишь одного элемента графической структуры, то впоследствии в результате упражнения такая структура процесса коренным образом меняется и письмо превращается в единую «кинетическую мелодию», не требующую специального припоминания зрительного образа изолированной буквы или отдельных двигательных импульсов для выполнения каждого штриха. Аналогичным образом развиваются и другие высшие психологические процессы.
Естественно, что в процессе такого развития меняется не только функциональная структура процесса, но и его мозговая локализация: участие слуховых и зрительных зон коры, обязательное на ранних этапах формирования некоторой деятельности, перестает быть необходимым на его поздних этапах, и та же деятельность начинает опираться на иную систему совместно работающих зон (А. Р.Лурия, Э. Г. Симерницкая, Б.Тыбулевич, 1970).
78
Развитие высших психических функций в онтогенезе имеет еще одну особенность, имеющую решающее значение для функциональной организации их в коре головного мозга.
Как показал в свое время Л.С.Выготский (1960), в процессе онтогенеза меняется не только структура высших психических процессов, но и их отношение друг к другу, иначе говоря, их «межфункциональная организация». Если на первых этапах развития сложная психическая деятельность имеет более элементарную основу и зависит от основной, «базальной», функции, то на дальнейших этапах развития она не только приобретает более сложную структуру, но и начинает осуществляться при ближайшем участии более высоких по своему строению форм деятельности.
Так, если маленький ребенок мыслит, опираясь на наглядные образы восприятия и памяти, иначе говоря, мыслит припоминая, то на более поздних этапах — юношеского или зрелого возраста — отвлеченное мышление с его операциями отвлечения и обобщения развивается настолько, что даже такие относительно простые процессы, как восприятие и память, превращаются в сложные формы познавательного анализа и синтеза, и человек теперь уже воспринимает и припоминает, размышляя.
Изменение отношений между основными психологическими процессами не может оставить неизменным соотношение основных систем мозговой коры, необходимых для осуществления этих процессов. Поэтому, если в раннем возрасте поражение какой-нибудь зоны коры, обеспечивающей относительно элементарные основы психической деятельности (например, зрительных отделов коры головного мозга), неизбежно вызывает в виде вторичного, системного эффекта неразвитие более высоких, надстроенных над ней образований, то у зрелого человека, у которого сложные системы не только уже сформировались, но и стали оказывать решающее влияние на организацию более простых форм деятельности, поражение низших зон уже не имеет такого значения, которое оно имело на ранних этапах развития, и, наоборот, поражение высших зон приводит к распаду элементарных функций, приобретших сложное строение и интимно зависящих от более высокоорганизованных форм деятельности.
Сформулированное Л.С.Выготским правило, согласно которому поражение определенной области мозга в раннем детстве системно влияет на более высокие зоны коры, надстраивающиеся над ними, в то время как поражение той же области в зрелом возрасте влияет на более низкие зоны коры, которые теперь от них зависят, является одним из фундаментальных положений, внесенных в учение о динамической локализации высших
79
психических функций отечественной психологической наукой. В качестве иллюстрации его укажем, что поражение вторичных отделов зрительной коры в раннем детстве может привести к системному недоразвитию высших процессов, связанных с наглядным мышлением, в то время как поражение этих же зон в зрелом возрасте может вызвать лишь частные дефекты зрительного анализа и синтеза, оставив сохранными уже сформировавшиеся раньше более сложные формы мышления. Сказанное о системном строении высших психологических процессов заставляет нас коренным образом пересмотреть классические представления о локализации их в коре головного мозга.
Свою основную задачу теперь мы видим в том, чтобы, тщательно проанализировав совместно работающие зоны мозга, обеспечивающие выполнение сложных форм психической деятельности, выяснить место каждой из этих зон в функциональной системе и то, как меняется соотношение этих совместно работающих отделов мозга при осуществлении психической деятельности на разных этапах развития.
Естественно, что такой подход в корне изменяет и практическую форму работы психолога. Прежде чем ответить на вопрос о том, каковы мозговые основы того или иного психического процесса человека, необходимо тщательно изучить строение того психологического процесса, мозговую организацию которого мы хотим установить, и выделить в нем те звенья, которые в той или иной степени могут быть отнесены к определенным системам мозга. Только такая работа по уточнению функциональной структуры изучаемого психологического процесса с выделением его составных компонентов и с дальнейшим анализом их «размещения» по системам головного мозга позволяет подойти к новому решению старого вопроса о локализации психических функций в коре головного мозга.
Пересмотр понятия «симптом»
Классические исследования локализации психических функций в коре головного мозга, пользовавшиеся наблюдениями над изменением поведения после ограниченных (локальных) поражений мозга, исходили из упрощенных представлений, согласно которым нарушение той или иной психической функции (речи, письма, чтения, праксиса, гнозиса), наступающее в результате разрушения определенного участка головного мозга, является прямым доказательством того, что данная функция локализована в этом (теперь разрушенном) участке мозга.
80
Рассмотренные выше факты привели к коренному пересмотру этих слишком упрошенных представлений. Естественно, что нарушение общей чувствительности неизбежно указывает на поражение задней центральной извилины, равно как выпадение части зрительного поля — на поражение сетчатки, зрительных путей или зрительной коры. В этих случаях установить симптом — значит получить отчетливые данные о топической диагностике поражения и тем самым о локализации функции в нервном аппарате.
Совершенно иначе обстоит дело в случаях нарушения высших психических процессов. Если психическая деятельность является сложной функциональной системой, в осуществление которой вовлекается целый комплекс совместно работающих зон мозговой коры, то поражение каждой из этих зон может привести к распаду всей функциональной системы, и, таким образом, симптом (нарушение или выпадение той или иной функции) еще ничего не говорит о ее локализации.
Для того чтобы перейти от установления симптома к локализации соответствующей психической деятельности, нужно пройти длинный путь, важнейшим звеном которого является детальный психологический анализ структуры возникающего нарушения и выяснение тех ближайших причин, вследствие которых функциональная система распалась, или, иначе говоря, дать тщательную квалификацию наблюдаемого симптома.
Поясним нашу мысль на примере. В клинике локальных поражений мозга очень часто встречается симптом апраксии, заключающийся в том, что больной оказывается неспособным выполнить те или иные предметные действия. В классической неврологии этого было достаточно, чтобы заключить, что поражение локализуется в нижнетеменной области, считавшейся «центром сложного прак-сиса». Если апраксия носила характер затрудненного выполнения движения при отчетливо представляемой схеме, его поражение локализовалось кпереди от нижнетеменной области.
После исследований ряда физиологов (и в первую очередь выдающегося отечественного физиолога Н. А. Бернштейна) стало совершенно ясным, что всякое произвольное движение — и в еще большей мере предметное действие — представляет собой сложную функциональную систему, включающую в свой состав ряд условий, при отсутствии которых такое движение не может быть выполнено.
Для выполнения движения необходима прежде всего кинестетическая афферентация, иначе говоря, система кинестетических импульсов, которые «сообщают» мозгу о положении движущейся конечности, суставных сочленений и степени вязкости мышц. Если такие афферентные импульсы (прием и синтез которых обеспечивается общечувствительными отделами постцентральной коры) выпадают, движение теряет афферентную основу, и эффекторные импульсы, идущие от коры головного мозга к мышцам, становятся практически неуправляемыми. Как следствие, даже относительно тонкие поражения постцентральной области коры могут приве-
81
сти к своеобразной кинестетической апраксии, которая принимает характер нарушения тонких, дифференцированных движений, приводящего к невозможности обеспечить нужную позу руки, адекватную тому предметному действию, которое рука выполняет. Наличие кинестетической афферентации важное, однако недостаточное условие выполнения соответствующего движения.
Всякое движение — будь то передвижение в пространстве, попадание в цель или действие с предметом — всегда осуществляется в системе пространственных координат и необходимо нуждается в синтезе зрительно-пространственных афферентации, что обеспечивается участием третичных, теменно-затылочных, отделов коры, куда стекаются импульсы от зрительного, вестибулярного и кожно-кинестетического анализаторов.
Если эта область коры поражается и пространственный синтез распадается, с необходимостью возникает нарушение сложно построенного движения. Однако апраксия, возникающая в этих случаях, носит совсем иной характер и проявляется прежде всего в невозможности придать действующей руке нужное положение в пространстве; так, например, больной начинает испытывать затруднения, застилая кровать, и часто вместо того чтобы разместить одеяло вдоль кровати размещает его поперек, он не может сохранить правильное направление ложки, которую он держит, придавая ей часто не горизонтальное, а вертикальное положение, и т.д.
Очевидно, что такая пространственная апраксия резко отличается от описанной выше кинестетической апраксии как по структуре действия, так и по механизмам и лежащей в ее основе локализации дефекта.
Перечисленных условий, однако, все еще недостаточно для осуществления полноценного движения или действия. Каждое действие всегда состоит из цепи последовательно развертывающихся движений, каждое звено которой после реализации должно быть денервировано с тем, чтобы уступить место следующему звену. На начальных стадиях формирования эта цепь двигательных звеньев носит развернутый характер, и каждое двигательное звено требует специального изолированного импульса. С формированием двигательного навыка цепь изолированных импульсов редуцируется и сложное движение начинает осуществляться как единая «кинетическая мелодия».
Существенным является тот факт, что кинетическая организация движений обеспечивается как базальными двигательными узлами (на ранних этапах формирования элементарных двигательных синергии), так и премоторными отделами коры (на поздних этапах формирования сложных двигательных навыков). Поэтому при поражении этих отделов коры также возникает апраксия, но на этот раз она носит характер кинетической апраксии, проявляясь в невозможности
82
синтезировать двигательные звенья в одну плавную мелодию, в задержках денервации ранее выполненного звена движения и затрудненности перехода от одного двигательного звена к другому. Естественно, что структура кинетической апраксии существенно отличается от ранее описанных форм нарушения праксиса, равно как и локализация соответствующих симптомов.
Нам осталось указать на последнее условие правильности выполнения движения.
Всякое движение направлено на известную цель и реализует определенную двигательную задачу. На уровне элементарно построенного инстинктивного поведения двигательные задачи диктуются врожденными программами; на уровне сложного, прижизненно сформированного, сознательного действия они определяются намерениями, которые в свою очередь сформированы при ближайшем участии речи, регулирующей поведение человека (А. Р. Лурия, 1961). Как было показано специальными исследованиями (А. Р. Лурия, 1962, 1963, 1966, 1969; А. Р.Лурия и Е.Д.Хомская, 1966), формирование сложных намерений, осуществляющееся посредством регулирующей речи, протекает при ближайшем участии лобных долей мозга. Поэтому массивные поражения лобных долей также могут привести к апраксии, но эта апраксия целевого действия коренным образом отличается от ранее описанных форм. Как правило, она сводится к невозможности подчинить свои движения сформированному в речи намерению, к распаду сложно организованных программ и к замене осмысленного, целенаправленного действия эхопраксическим повторением движений исследующего или инертными стереотипами, теряющими осмысленный, целенаправленный характер. Мы не будем сейчас останавливаться на этом типе нарушений праксиса, поскольку далее он будет предметом специального обсуждения.
Приведенные факты позволяют нам прийти к важному заключению. Симптом нарушения праксиса (апраксия) является признаком локального поражения мозга; однако сам по себе этот симптом еще не говорит о локализации очага. Произвольное движение (праксис) является сложнейшей функциональной системой, которая базируется на сохранности ряда факторов и опирается на совместную работу целого комплекса зон мозговой коры и подкорковых образований, причем каждая из этих зон вносит свой вклад в осуществление движения. Поэтому сложное предметное движение может нарушаться при поражении различных зон коры (или подкорковых образований), однако каждый раз оно будет нарушаться по-разному.
Ближайшая задача исследователей сводится к тому, чтобы, изучив структуру наблюдаемого дефекта, квалифицировать симптом. Только такая работа, приводящая к выделению основного фактора, стоящего за наблюдаемым симптомом, позволяет прийти к заключению о локализации очага, лежащего в основе дефекта.
83
Таким образом, понятия «локализация очага» и «локализация функции» не совпадают, и для того чтобы использовать метод локальных поражений мозга для заключения о локализации функции (или, точнее, о мозговой организации функциональной системы), необходимо осуществить структурный анализ синдрома. Это и есть основной путь нейропсихологического исследования.
СИНДРОМНЫЙ АНАЛИЗ И СИСТЕМНАЯ ОРГАНИЗАЦИЯ ПСИХИЧЕСКИХ ПРОЦЕССОВ
Квалификация симптома является лишь первым этапом анализа мозговой организации психических процессов. Для того чтобы данные о локальной патологии мозга позволили прийти к надежным выводам относительно как структуры психических процессов, так и их локализации в коре головного мозга человека, необходимо перейти от квалификации единичного симптома к описанию целого симптомокомплекса, или, как это принято называть, к синдромному анализу изменений поведения, наступающих при локальном поражении мозга.
Как мы уже сказали, любая психическая деятельность человека является сложной функциональной системой, реализация которой обеспечивается целым комплексом совместно работающих аппаратов мозга, каждый из которых вносит свой вклад в обеспечение этой функциональной системы. Это практически означает, что функциональная система в целом может нарушаться при поражении большого числа зон, причем при различных по локализации поражениях она нарушается по-разному. Последнее положение связано с тем, что каждая зона мозга, участвующая в обеспечении функциональной системы, ответственна за свой фактор, и его устранение приводит к тому, что нормальное осуществление функции становится невозможным. Приведенный выше пример построения произвольного движения и видов его нарушения при локальных поражениях мозга отчетливо это иллюстрирует.
Описанные закономерности построения и распада функциональных систем позволяют нейропсихологу выяснить, какие именно факторы включает в свой состав сложная психическая деятельность и какие именно участки мозга составляют ее нервную основу.
Оба эти вопроса могут быть решены только при сопоставлении всех симптомов, которые отмечаются при наличии одного, строго локализованного очага в коре головного мозга (или в подкорковых образованиях), с одной стороны, и тщательного анализа характера нарушений данной системы при различных по локализации мозговых поражениях — с другой.
Остановимся на иллюстрации этого положения.
84
Как уже было сказано ранее, для успешного осуществления сложного движения нужна его четкая пространственная организация, иначе говоря, четкое построение движения в системе пространственных координат. Это условие обеспечивается третичными (зрительно-кинестетически-вестибулярными) отделами теменно-затылочной коры, несоблюдение этого условия приводит к распаду пространственно-организованного движения. Возникает, однако, естественный вопрос: какие другие виды психической деятельности нарушаются при поражении тех же теменно-затылочных отделов мозга, обеспечивающих функцию пространственной организации процессов? Если мы получим ответ на этот вопрос и если мы сможем выделить одну группу процессов, страдающих при такой локализации очага, и другую группу процессов, которая при данном патологическом очаге остается сохранной, мы значительно приблизимся к выяснению того, в какие именно виды психической деятельности входит пространственный фактор, имеющий прямую связь с указанными теменно-затылочными отделами мозговой коры.
Факты, показывающие, что любой ограниченный корковый очаг поражения действительно нарушает протекание одних психических процессов, оставляя другие процессы в сохранности (явление, названное американским нейропсихологом Г.Л.Тэйбером принципом двойной диссоциации функций), в изобилии содержатся в материалах нейропсихологических исследований.
Так, ограниченный очаг в теменно-затылочной (или нижнетеменной) области левого полушария, приводящий к нарушению пространственной организации восприятия и движения, неизбежно вызывает и другие симптомы: такие больные, как правило, не могут ориентироваться в положении стрелок на часах или в координатах на географической карте; они не могут ориентироваться в плане клинического отделения, где они находятся; не могут решить даже относительно несложные арифметические примеры, смешивая, например, при вычитании из двузначного числа с переходом через десяток направление операций (решая пример 31 — 7, они правильно выполняют первый этап — от 30 отнимают 7 и получают 23, но затем не знают, направо или налево надо отложить оставшуюся единицу и какой окончательный ответ — «22» или «24» является правильным); наконец, они начинают испытывать серьезные затруднения в понимании ряда грамматических структур, включающих в свой состав известные логические отношения (например, «брат отца» и «отец брата», «весна после лета» или «лето после весны»), в то время как понимание других, более простых грамматических структур остается сохранным.
Однако указанный очаг не приводит к нарушениям таких процессов, как плавная речь, узнавание или воспроизведение музыкальных мелодий, смена последовательных элементов движения и т.д.
85
Все это показывает, что первая группа отмеченных выше процессов включает в свой состав «пространственный» фактор, в то время как вторая группа процессов этого фактора не включает и поэтому остается сохранной при поражении теменно-затылочных отделов коры.
Совершенно обратная картина наблюдается при локальных поражениях височной (слуховой) области коры. Поражения с такой локализацией, как мы увидим далее, приводят к нарушению организации слухового восприятия и делают невозможной организацию звуковых сигналов в последовательную («сукцессивную») структуру. Именно в силу этого больные с такими поражениями оказываются не в состоянии четко воспринимать обращенную к ним речь и удерживать ее следы; плавная избирательная речь, как и слухоречевая память, оказывается у них серьезно нарушенной. Однако такие функции, как ориентировка в пространстве, пространственная организация движений, счетные операции, понимание определенных логико-грамматических отношений, остаются у них, как правило, сохранными.
Все это с полной отчетливостью показывает, что тщательный нейропсихологический анализ синдрома и той двойной диссоциации, которая возникает при локальных поражениях мозга, позволяет приблизиться к структурному анализу самих психологических процессов и выделить те факторы, которые входят в одни группы психических процессов и не входят в другие.
Как мы увидим далее, это позволяет вплотную подойти к решению вопроса о внутреннем составе психологических процессов, который оставался неразрешимым в условиях обычного психологического исследования, и, таким образом, разделить, казалось бы, сходные психологические процессы и сблизить, казалось бы, различные формы психической деятельности.
Проиллюстрируем это положение двумя примерами.
Для непредвзятого наблюдателя музыкальный и речевой слух могут казаться двумя вариантами одного и того же психологического процесса. Однако наблюдения над больными с ограниченными очаговыми поражениями мозга показывают, что разрушение определенных участков левой височной области приводит к выраженному нарушению речевого слуха (делая различение близких звуков речи совершенно недоступным), но сохраняет музыкальный слух неповрежденным. В наших публикациях есть описание одного выдающегося композитора, который после кровоизлияния в левую височную область перестал различать звуки речи и понимать обращенную к нему речь, но продолжал создавать блестящие музыкальные произведения (А. Р. Лурия, Л. С. Цветкова, Д. С. Футер, 1965).
Это означает, что столь близкие, казалось бы, психические процессы, как музыкальный и речевой слух, не только включают в свой состав разные факторы, но и опираются на работу различных мозговых зон.
86
Нейропсихологии известны также примеры, показывающие внутреннюю близость, казалось бы, полностью различных психологических процессов.
Вряд ли кто-нибудь сразу согласится с тем, что столь различные психологические процессы, как ориентировка в пространстве, счет и понимание сложных логико-грамматических структур, имеют принципиально общие звенья, которые позволяют объединить их в одну группу.
Однако поражение теменно-затылочных (нижнетеменных) отделов левого полушария почти неизбежно приводит к нарушению всех этих процессов, и больной с подобной локализацией очага не только испытывает заметные трудности в пространственной ориентировке, но и обнаруживает грубейшие дефекты в счете и в понимании сложных логико-грамматических структур.
Это показывает, что все указанные, казалось бы, столь различные функции включают общий фактор, и выделение этих общих факторов способствует гораздо более глубокому анализу структуры психологических процессов.
Нетрудно увидеть, что синдромный анализ позволяет не только уточнить вопрос о мозговой организации сложных психических процессов, но и вплотную подойти к их внутреннему строению.
Итак, каждая сознательная психическая деятельность представляет собой сложную функциональную систему, которая может нарушаться в разных звеньях и страдать при различных по локализации поражениях головного мозга (но по-разному); в ее состав включены различные факторы, изучение которых открывает новые пути для нейрофизиологического анализа внутреннего строения психических процессов.
Наблюдения над изменениями психических процессов, возникающими при локальных поражениях мозга, могут действительно стать одним из наиболее существенных источников наших знаний о мозговой организации психической деятельности. Однако правильное использование этого метода возможно лишь при условии отказа от концепции прямой локализации психических процессов в коре головного мозга и замены этой классической задачи другой, требующей анализа того, как именно меняется психическая деятельность при различных локальных поражениях мозга и какие именно факторы вносит каждый из аппаратов головного мозга в построение сложных форм психической деятельности.
Эта новая задача определяет генеральный путь нейропсихологии как учения о мозговой организации психических процессов человека.
Глава III
ТРИ ОСНОВНЫХ ФУНКЦИОНАЛЬНЫХ БЛОКА МОЗГА
Мы уже говорили, что психические процессы человека являются сложными функциональными системами и что они не локализованы в узких, ограниченных участках мозга, а осуществляются при участии сложных комплексов совместно работающих мозговых аппаратов, каждый из которых вносит свой вклад в организацию этой функциональной системы. Вот почему становится необходимым выяснить, из каких основных функциональных единиц состоит мозг человека, как построена и какую роль играет каждая из них в осуществлении сложных форм психической деятельности.
Можно выделить три основных функциональных блока, или три основных аппарата мозга, участие которых необходимо для осуществления любого вида психической деятельности. С некоторым приближением к истине их можно обозначить как: 1) блок, обеспечивающий регуляцию тонуса и бодрствования; 2) блок получения, переработки и хранения информации, поступающей из внешнего мира; 3) блок программирования, регуляции и контроля психической деятельности.
Каждый из этих основных блоков имеет иерархическое строение и состоит по крайней мере из надстроенных друг над другом корковых зон трех типов: первичных (или проекционных), куда поступают импульсы с периферии или откуда направляются импульсы на периферию, вторичных (или проекционно-ассоциативных), где происходит переработка получаемой информации или подготовка соответствующих программ, и, наконец, третичных (или зон перекрытия), которые являются наиболее поздно развивающимися аппаратами больших полушарий и которые у человека обеспечивают наиболее сложные формы психической деятельности, требующие совместного участия многих зон мозговой коры.
Рассмотрим строение и функциональные особенности каждого из этих блоков головного мозга в отдельности.
1 БЛОК РЕГУЛЯЦИИ ТОНУСА И БОДРСТВОВАНИЯ
Для того чтобы обеспечивалось полноценное протекание психических процессов, человек должен находиться в состоянии бодрствования. Известно, что только в оптимальных условиях бодрствования человек может принимать и перерабатывать информацию, вызывать в памяти нужные избирательные системы связей, программировать свою деятельность и осуществлять контроль за протеканием своих психических процессов, корригируя ошибки и сохраняя направленность своей деятельности.
Хорошо известно, что в состоянии сна четкая регуляция психических процессов невозможна, всплывающие воспоминания и ассоциации приобретают неорганизованный характер, и направленное избирательное (селективное) выполнение психической деятельности становится невозможным.
О том, что для осуществления организованной, целенаправленной деятельности необходимо поддерживать оптимальный тонус коры, говорил еще И.П.Павлов, гипотетически утверждавший, что если бы мы могли видеть, как распространяется возбуждение по коре бодрствующего животного (или человека), мы наблюдали бы «светлое пятно», перемещающееся по коре мозга по мере перехода от одной деятельности к другой и олицетворяющее пункт оптимального возбуждения.
Развитие электрофизиологической техники позволило увидеть это «пятно» оптимального возбуждения: с помощью специального прибора — «топоскопа» М.Н.Ливанова (1962), дающего возможность одновременно регистрировать электрическую активность в 50—100 пунктах коры головного мозга, — можно наблюдать, как в коре мозга бодрствующего животного действительно возникает «пятно» оптимального возбуждения, как оно передвигается при переходе животного из одного состояния в другое и как в патологическом состоянии постепенно теряет свою подвижность, становится инертным или совсем угасает.
И. П. Павлов не только указал на необходимость оптимального состояния мозговой коры для осуществления организованной деятельности, но и открыл основные нейродинамические законы возникновения такого оптимального состояния. Как было показано многочисленными исследованиями павловской школы, процессы возбуждения и торможения, протекающие в бодрствующей коре, подчиняются закону силы, характеризуются определенной концентрированностью, уравновешенностью и подвижностью.
Эти основные законы нейродинамики неприложимы к состояниям сна или утомления. Это является результатом того, что в так называемых «тормозных», или «фазовых», состояниях тонус коры снижается и, как следствие, нарушается закон силы: слабые раздражители уравниваются с сильными по интенсивности вызываемых ими ответов («уравнительная фаза») или даже превосходят их, вызывая более интенсивные реакции, чем те, которые вызываются сильными раздражителями («парадоксальная фаза»), в отдельных случаях реакции сохраняются только в ответ на слабые
89
стимулы, в то время как сильные раздражители вообще перестают вызывать какие-либо ответы («ультрапарадоксальная фаза»). Кроме того, по мере снижения тонуса коры нарушается нормальное соотношение возбудительных и тормозных процессов и та подвижность, которая необходима для протекания нормальной психической деятельности. Все это указывает на то, какое решающее значение имеет наличие оптимального тонуса коры для организованного протекания психической деятельности.
Возникает, однако, вопрос: какие аппараты мозга обеспечивают поддержание оптимального тонуса коры, о котором мы только что говорили? Какие участки мозга регулируют и изменяют тонус коры, сохраняя его на нужное время и повышая его, когда в этом возникает необходимость?
Одним из наиболее важных в этом плане открытий было установление того факта, что аппараты, обеспечивающие и регулирующие тонус коры, могут находиться не в самой коре, а в лежащих ниже стволовых и подкорковых отделах мозга и что эти аппараты находятся в двойных отношениях с корой, тонизируя ее и в то же время испытывая ее регулирующее влияние.
В 1949 г. два выдающихся исследователя — Мэгун и Моруцци — обнаружили, что в стволовых отделах головного мозга находится особое нервное образование, которое как по своему морфологическому строению, так и по своим функциональным свойствам приспособлено к тому, чтобы осуществлять роль механизма, регулирующего состояния мозговой коры, т. е. способно изменять ее тонус и обеспечивать ее бодрствование.
Это образование построено по типу нервной сети, в которую вкраплены тела нервных клеток, соединяющиеся друг с другом короткими отростками. По сети этого образования, названного ретикулярной формацией, возбуждение распространяется не отдельными, изолированными импульсами, не по закону «все или ничего», а градуально, постепенно меняя свой уровень и, таким образом, модулируя состояние всего нервного аппарата (рис. 34).
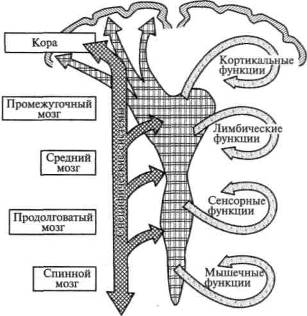
Рис. 34. Схема активирующей ретикулярной формации
Одни из волокон ретикулярной формации направляются вверх, оканчиваясь в расположенных выше нервных образованиях — зрительном бугре, хвостатом теле, древней коре и, наконец, в тех образованиях новой коры, роль которых в организации сложной психической деятельности была отмечена в предьщущих главах. Эти образования были названы восходящей ретикулярной системой. Как обнаружили последующие наблюдения, она играет решающую роль в активации коры, в регуляции ее активности.
Другие волокна ретикулярной формации имеют обратное направление: они начинаются от более высоко расположенных нервных образований — новой и древней коры, хвостатого тела и ядер зрительного бугра — и направляются к расположенным ниже структурам среднего мозга, гипоталамуса и мозгового ствола. Эти обра-
90
зования получили название нисходящей ретикулярной системы. Они, как было установлено дальнейшими наблюдениями, ставят нижележащие образования под контроль тех программ, которые возникают в коре головного мозга и для выполнения которых требуется модификация и модуляция состояний бодрствования.
Оба раздела ретикулярной формации составляют единую вертикально расположенную функциональную систему, единый саморегулирующийся аппарат, построенный по принципу рефлекторного круга, который может обеспечивать изменение тонуса коры, но который вместе с тем сам находится под регулирующим влиянием тех изменений, которые наступают в коре головного мозга. Это аппарат пластичного приспособления к условиям среды в процессе активной деятельности.
С открытием ретикулярной формации в нейрофизиологию был введен фактически новый принцип — вертикальной организации всех структур мозга — и завершился длительный период, когда интерес ученых, пытавшихся найти нервные механизмы психических процессов, был сосредоточен лишь на аппаратах коры головного мозга, работа которой рассматривалась как не зависящая от нижележащих, глубоких образований. Ретикулярная формация — первый функциональный блок головного мозга — аппарат, обеспечивающий регуляцию тонуса коры и состояний бодрствования, аппарат, позволяющий регулировать эти состояния соответственно поставленным перед организмом задачам.
91
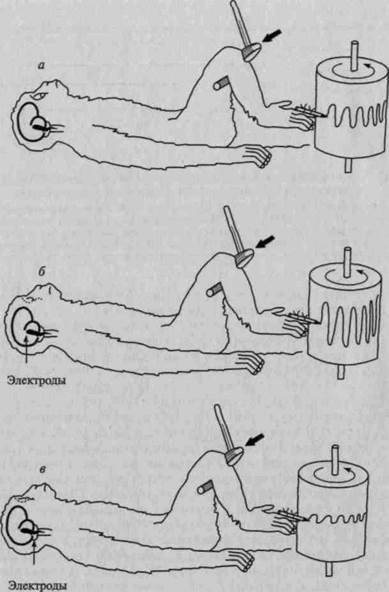
Рис. 37. Активирующая роль раздражения ретикулярной формации на двигательную сферу (коленный рефлекс):
а — до раздражения ретикулярной формации; б — во время раздражения ретикулярной формации; в — после раздражения ретикулярной формации
(по Френчу)
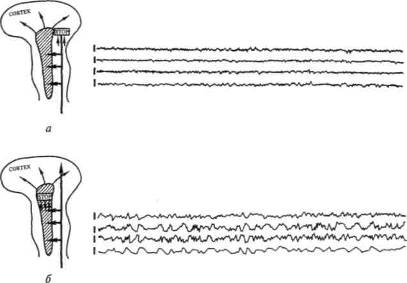
Рис. 38. Развитие сна в результате перерезки путей активирующей ретикулярной формации (по Линдсли): а — активное состояние мозга в результате сохранения активирующих влияний ретикулярной формации на кору больших полушарий; электроэнцефалограмма бодрствования; б — состояние сна вследствие перерезки верхних отделов стволовой ретикулярной формации и прекращения активирующих влияний; электроэнцефалограмма сна
Регулирующая тонус коры, модулирующая ее состояние, функция ретикулярной формации была подтверждена большим количеством экспериментов, проведенных такими исследователями, как Моруцци и Мэгун (1949), Линдсли и др. (1949, 1960, 1961), Бремер (1954, 1957), Джаспер (1954, 1957, 1963), Френч и др. (1955), Сегундо и др. (1955), Жувэ и др. (1957, 1959), Наута (1964, 1968), Прибрам (1966, 1967, 1969, 1971). Эти исследования показали, что раздражение ретикулярной формации (в области среднего мозга, задней части гипоталамуса и примыкающих к ним субталамических структур) вызывает реакцию пробуждения (рис. 35), повышает возбудимость, обостряет (снижая абсолютные и различительные пороги) чувствительность (Линдсли, 1951, 1958, 1960; и др.) (рис. 36) и оказывает тем самым общее активирующее влияние на кору головного мозга (рис. 37). Существенным оказался и тот факт, что поражение этих структур приводит к резкому снижению тонуса коры, к появлению состояния сна с картиной синхронизации в ЭЭГ (рис. 38), а иногда и к коматозному состоянию. У животных в этих случаях реакция arousal отсутствует даже в ответ на сильные болевые раздражения (Линдсли и др., 1949; Френч, Мэгун, 1952; Френч, 1952; С. П.Нарикашвили, 1961, 1963, 1968).

Рис. 35. Активирующее влияние раздражения ретикулярной формации на кору мозга, вызывающее реакцию пробуждения (по Френчу). Кошка пробуждается от действия звонка; возбуждения, возникающие в ретикулярной формации, распространяются на слуховую зону коры и ведут к пробуждению. Соответственно меняются и волны электроэнцефалограммы. Ретикулярная формация интегрирует мозговую активность и приводит к общей организованной реакции кошки
Таким образом, ретикулярная формация ствола является одним из мощных механизмов тонизации коры головного мозга, регулирующих функциональное состояние мозга, или уровень бодрствования.

Рис. 36. Активирующее влияние раздражения ретикулярной формации на повышение чувствительности. Различение двух вспышек:
а — до раздражения ретикулярной формации; б — во время раздражения; в —
через 0 — 10 с; г — через 10 — 12 с после раздражения. 1-я линия — вспышки;
2-я — реакции зрительного тракта; 3-я — реакции наружного коленчатого
тела; 4-я и 5-я — реакции зрительной коры (по Линдсли). Обратить внимание
на двойную реакцию, появляющуюся в зрительной коре после раздражения
93
Эксперименты, проведенные рядом авторов (Жувэ, 1961; Эрнандес-Пеон, 1965, 1969; С. П. Нарикашвили и др., 1962, 1963, 1963, 1968; Загер, 1968), показали, что наряду с активирующими существуют и тормозящие разделы ретикулярной формации. Вот почему раздражение одних ядер ретикулярной формации неизменно приводит к активации животного, в то время как раздражение других ядер ведет к возникновению характерных для сна изменений в электрической активности коры и к развитию сна.
Этот факт в равной степени относится и к мозгу животного, и к мозгу человека. Именно поэтому, раздражая во время нейрохирургических операций стенки третьего желудочка, выдающийся отечественный хирург Н. Н. Бурденко мог искусственно вызывать на операционном столе сон у больного.
Участие образований первого блока головного мозга верхнего ствола и ретикулярной формации в регуляции состояний бодрствования не вызывает сомнений. Мы еще раз убедимся в этом, когда перейдем к изучению тех нарушений, которые возникают в психических процессах человека при поражении этих отделов мозга.
95
Активирующая ретикулярная формация, являющаяся важнейшей частью первого функционального блока мозга, с самого начала была названа неспецифической; это коренным образом отличало ее от других, специфических (сенсорных и двигательных) систем мозговой коры. Считалось, что ее активирующее и тормозное действие равномерно затрагивает как все сенсорные, так и все двигательные функции организма и что ее функцией является лишь регуляция состояний сна и бодрствования — того неспецифического фона, на котором протекают различные виды деятельности.
Это утверждение, которое было одним из исходных для классиков нейрофизиологии, впервые описавших активирующую систему мозга, нельзя, однако, признать полностью правильным. Как показали дальнейшие наблюдения, ретикулярная система мозга имеет определенные черты дифференцированности, или специфичности, как по своим анатомическим характеристикам (Бродал, 1957; М. Шейбел и А. Шейбел, 1958), так и по источникам активации и формам работы, однако эта специфичность не имеет ничего общего с «модальностью» органов чувств (анализаторов) (П. К.Анохин, 1958, 1961, 1963; Иошии, 1966, 1969).
Известно, что нервная система всегда находится в состоянии определенной активности и что наличие некоторого тонуса обязательно для любого проявления жизнедеятельности. Можно выделить по крайней мере два основных источника активации; действие каждого из них опосредствуется активирующей ретикулярной формацией, однако, что существенно, ее различными частями. В этом и состоит дифференцированность, или специфичность, функциональной организации «неспецифической активирующей системы». Первыми из источников активации являются обменные процессы организма, лежащие в основе гомеостаза (внутреннего равновесия организма) и инстинктивных процессов.
Обменные процессы (или, как иногда выражаются, «внутреннее хозяйство организма») в их наиболее простых формах связаны с дыхательными и пищеварительными процессами, с процессами сахарного и белкового обмена, с процессами внутренней секреции и т.д.; все они регулируются главным образом аппаратами гипоталамуса. Тесно связанные с гипоталамусом ретикулярные формации продолговатого мозга (бульбарная) и среднего мозга (мезэнцефально-гипоталамическая) играют значительную роль в этой наиболее простой, «витальной», форме активации.
Более сложные формы этого вида активации связаны с обменными процессами, которые организованы в определенные врожденные системы поведения; они широко известны как системы инстинктивного (или безусловнорефлекторного) пищевого и поло-
96
вого поведения. Общим для обоих видов является то, что и в том и в другом случае источником активации являются обменные (гуморальные) процессы организма; отличаются они, во-первых, уровнем сложности организации и, во-вторых, тем, что если формы первого вида, наиболее элементарные, вызывают лишь примитивные автоматические реакции, связанные с недостатком кислорода или необходимостью выделения запасных веществ из их органических депо при голодании, то вторые предполагают организацию сложных поведенческих систем, в результате действия которых удовлетворяются определенные потребности и восстанавливается равновесие «внутреннего хозяйства организма».
Естественно, что для того чтобы обеспечить сложные инстинктивные формы поведения, необходима весьма избирательная специфическая активация, и биологически-специфические формы этой пищевой или половой активации являются функцией более высоко расположенных образований мезэнцефальной, диэнцефальной и лимбической систем. Большое число опытов, проведенных за последнее время (здесь и далее имеется в виду конец XX в. - примеч. ред.) (Олдс, 1958; Мак-Лин, 1959, 1960; Н.Миллер, 1966), дают возможность убедиться в том, что в упомянутых образованиях мозгового ствола и древней коры имеются высокоспецифические ядра ретикулярной формации, раздражение которых приводит либо к активации, либо к блокированию различных форм инстинктивного поведения. На рис. 39 мы приводим схему расположения ядер, активирующих или блокирующих пищевое, половое и оборонительное поведение.
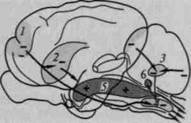
Рис. 39. Возбуждающее и тормозящее влияние раздражения ядер ретикулярной формации:
5—6 — облегчающие; 1, 2, 3 и 4 — тормозящие зоны ретикулярной формации ствола, а также связи, идущие к ней от коры (1) и от мозжечка (3)
Второй источник активации имеет совсем иное происхождении. Он связан с поступлением в организм раздражений из внешнего мира и приводит к возникновению совершенно иных форм активации, проявляющихся в виде ориентировочного рефлекса.
Человек живет в мире информации, и потребность в ней иногда оказывается у него не меньшей, чем потребность в органическом обмене веществ.
Известно, что человек, лишенный обычного притока информации — что имеет место в редких случаях выключения всех воспринимающих органов, — впадает в сон, из которого его может вывести только поступление какой-либо информации. Известно, что нормальный человек тяжело переносит ограничение контактов с внешним миром, и, как это наблюдал Хэбб (1955), достаточно поместить испытуемых в условия резкого ограничения притока возбуждений (сенсорной депривации), чтобы у них возникли психические нарушения, галлюцинации, в какой-то мере компенсирующие это ограничение. В аппаратах ретикулярной формации имеются специальные механизмы, обеспечивающие тоническую форму активации, источником которой является главным образом приток возбуждений из органов чувств. Этот источник обладает не меньшей интенсивностью, чем первый, только что упомянутый, источник активации. Однако тоническая форма активации, связанная с работой различных органов чувств, является лишь наиболее элементарным источником активации того типа, который мы описываем.
Человек живет в условиях постоянно меняющейся среды, и это требует обостренного состояния бодрствования. Обострением бодрствования сопровождается всякое изменение в окружающих условиях, появление любого (как неожиданного, так и ожидаемого) события. Такая мобилизация организма лежит в основе особого вида активности, которую Павлов называл ориентировочным рефлексом и которая является основой познавательной деятельности.
Одним из наиболее важных открытий последних десятилетий было обнаружение связи ориентировочного рефлекса, или реакции пробуждения (активации), с работой ретикулярной формации мозга (Моруцци, Мэгун, 1949; Г.В.Гершуни, 1949, 1955; Линдсли и др., 1949; Шарплес, Джаспер, 1956; Гасто, 1958; Е.Н.Соколов, 1958, 1964; О.С.Виноградова, 1961). Как показали исследования, ориентировочный рефлекс — сложное явление. Описаны тоническая и генерализованная формы реакции пробуждения, с одной стороны, и фазическая и локальная ее формы — с другой (Шарплес, Джаспер, 1956; Е.Н.Соколов, 1958, 1964; Линдсли и др., 1950; Гасто, 1958; Эди и др., 1960; О.С.Виноградова, 1961; Морелл, 1967). Упомянутые формы ориентировочного рефлекса связаны с различными структурами в пределах ретикулярной формации: тоническая и генерализованная формы — с нижними, а фазическая и локальная — с верхними отделами ствола, и прежде всего с неспецифической таламической системой.
Как показали микроэлектродные исследования, неспецифические ядра зрительного бугра, а также хвостатого тела и гиппокампа функционально тесно связаны с системой ориентировочного рефлекса (Джаспер, 1964; О.С.Виноградова, 1969, 1970; Н.Н.Данилова, 1966, 1969, 1970). Каждая реакция на «новизну» требует прежде всего сличения нового раздражителя с системой старых, уже появлявшихся ранее, раздражителей. Только такое сравнение («компарация») может установить, является ли
98
данный раздражитель действительно новым и требует ориентировочного рефлекса, или же уже знакомым, безразличным, не требующим специальной мобилизации организма. Только такой механизм может обеспечивать процесс «привыкания», когда многократно повторяющийся раздражитель теряет свою новизну и необходимость специальной мобилизации организма при его появлении исчезает. Очевидно, что в этом звене механизм ориентировочного рефлекса тесно связан с механизмами памяти, и именно связь обоих процессов обеспечивает компарацию сигналов, которая является одним из важнейших условий этого вида активации.
Важнейшим открытием последних лет было указание на тот факт, что значительная часть нейронов гиппокампа и хвостатого тела, не имеющих модально-специфических функций, осуществляют функцию компарации сигналов, реагируя на появление новых раздражителей и прекращая активность по мере привыкания к ним (О.С.Виноградова, 1969, 1970).
Активирующая и тормозящая, иначе говоря модулирующая, функция нейронов гиппокампа и хвостатого тела оказалась, как это стало ясным лишь в самые последние годы, основным источником регуляции тонических состояний мозговой коры, которые связаны с наиболее сложными формами ориентировочного рефлекса, носящими уже не врожденный, а прижизненно возникающий, или условнорефлекторный, характер.
Далее (см. ч. вторая, гл. I) мы увидим, как влияет нарушение нормальной функции этих областей мозга на протекание психических процессов человека.
Остановимся в самых общих чертах на третьем и, пожалуй, наиболее интересном для нас источнике активации, в котором описанный только что функциональный блок мозга принимает самое непосредственное участие.
Источниками активности человека являются не только обменные процессы или непосредственный приток информации, вызывающий ориентировочный рефлекс. Значительная часть активности человека обусловлена намерениями и планами, перспективами и программами, которые формируются в процессе его сознательной жизни, являются социальными по своему заказу и осуществляются при ближайшем участии сначала внешней и потом его внутренней речи.
Всякий сформулированный в речи замысел преследует некоторую цель и вызывает целую программу действий, направленных на достижение этой цели. Достижение цели прекращает активность. Было бы, однако, неправильным считать возникновение намерений и формулировку целей чисто интеллектуальным актом. Осуществление замысла или достижение цели требует определенной энергии и может быть обеспечено лишь при наличии некоторого уровня активности.
99
Источники такой активности оставались долгое время неизвестными, и только исследования последних лет позволили сделать существенный шаг к тому, чтобы ответ на этот вопрос стал яснее.
Исследования, которые мы имеем в виду, заставляют отвергнуть старые предположения о том, что эти источники следует искать только во внутрикортикальных связях; они убедительно показывают, что в поиске механизмов наиболее высоких форм организации активности следует опираться на все тот же вертикальный принцип строения функциональных систем мозга и, следовательно, обратиться к связям между высшими отделами коры и нижележащей ретикулярной формацией.
До сих пор, обсуждая механизмы работы первого функционального блока, мы рассматривали только восходящие связи активирующей ретикулярной системы, однако мы упоминали, что наряду с ними существуют и нисходящие связи коры и нижележащих образований; именно эти связи и осуществляют регулирующее влияние мозговой коры на нижележащие стволовые образования и являются механизмом, с помощью которого возникшие в коре функциональные узоры возбуждения вовлекают аппараты ретикулярной формации древнего мозга и получают энергетический заряд.
Нисходящие аппараты ретикулярной формации исследованы значительно меньше, чем восходящие связи. Однако благодаря целой серии работ (Френч и др., 1955; Сегундо и др., 1955; Галамбос, 1956; О.С.Адрианов, 1963; Мэгун, 1963; С.П.Нарикашвили и др., 1961, 1962, 1963, 1968) выяснилось, что посредством кортико-ретикулярных путей раздражение отдельных участков коры может вызывать генерализованную реакцию пробуждения (Брезье, 1960; Галамбос и др., 1960), оказывать облегчающее влияние на спинальные рефлексы, изменять — через систему гамма-афферентов — возбудимость мышц, повышать возбудимость кохлеарного аппарата (Эрнандес-Пеон, 1956; С. П. Нарикашви-ли, 1963), понижать пороги различительной чувствительности (Жувэ, Эрнандес-Пеон, 1957; Линдсли, 1951, 1957, 1960).
Таким образом, как морфологическими, так и морфофизио-логическими исследованиями с достаточной надежностью установлено, что наряду со специфическими сенсорными и двигательными функциями, на которых мы еще остановимся, кора головного мозга имеет и неспецифические активирующие функции и что раздражение определенных участков коры может оказывать как активирующие, так и тормозящие влияния на нижележащие нервные образования (Жувэ, 1961; Бузер и др., 1961; С.П.Нарикашвили, 1963, 1968; О.Загер, 1968; Эрнандес-Пеон, 1966, 1969; Р.А.Дуринян и др., 1968). Оказалось далее, что нисходящие волокна активирующей (и тормозящей) ретикулярной системы имеют достаточно дифференцированную корковую организацию; если связанные со специфическими путями пучки волокон (повышающих или по-
100
нижающих тонус сенсорных или двигательных аппаратов) исходят из первичных (и частично из вторичных) зон коры, то волокна, опосредствующие более общие активирующие влияния на ретикулярную формацию ствола, исходят прежде всего из лобных отделов хоры (Френч и др., 1955; Сегундо и др., 1955; На-ута, 1964, 1968; Прибрам, 1959, 1960, 1964, 1971; О.Загер, 1968; Е.Д.Хомская, 1969, 1972) (рис. 40). Нисходящие волокна, идущие преимущественно от префронтальной (орбитальной и медиальной) коры, адресуются к ядрам зрительного бугра и нижележащих стволовых образований и являются тем аппаратом, посредством которого высшие отделы мозговой коры, непосредственно участвующие в формировании намерений и планов, управляют работой нижележащих аппаратов ретикулярной формации таламуса и ствола, тем самым модулируя их работу и обеспечивая наиболее сложные формы сознательной деятельности.
Рис. 40. Схема соотношения корковых аппаратов с образованиями
ствола посредством ретикулярной формации (по Мэгуну):
а — нисходящие; б — восходящие пути
Все это показывает, во-первых, что аппараты первого функционального блока не только тонизируют кору, но и сами испытывают ее дифференцирующее влияние, и, во-вторых, что первый функциональный блок мозга работает в тесной связи с высшими отделами коры.
2 БЛОК ПРИЕМА, ПЕРЕРАБОТКИ И ХРАНЕНИЯ ИНФОРМАЦИИ
В предыдущем параграфе мы говорили об аппаратах первого функционального блока головного мозга, который играет важную роль в регуляции состояний активности коры и уровня бодрствования. Этот блок построен по типу неспецифической нервной сети, которая осу-
101
ществляет свою функцию путем постепенного, градуального изменения состояний и не имеет непосредственного отношения ни к приему и переработке поступающей извне информации, ни к выработке намерений, планов и программ поведения. Этим первый функциональный блок мозга, расположенный в основном в пределах мозгового ствола, образований межуточного мозга и медиальных отделов новой коры, существенно отличается от аппаратов второго функционального блока мозга, основная функция которого заключается в приеме, переработке и хранении внешней информации.
Этот блок расположен в конвекситальных (наружных) отделах новой коры (неокортекса) и занимает ее задние отделы, включая в свой состав аппараты зрительной (затылочной), слуховой (височной) и общечувствительной (теменной) областей. По своему гистологическому строению он состоит из нейронов подкорки и мозговой коры. Эти нейроны в отличие от аппаратов первого блока работают не по принципу градуальных изменений, а по закону «все или ничего», принимая отдельные импульсы и передавая их на другие группы нейронов.
По своим функциональным особенностям аппараты этого блока приспособлены к приему экстероцептивных раздражений, приходящих в головной мозг от периферических рецепторов, к дроблению их на огромное число компонентов (иначе говоря, к анализу их на мельчайшие составляющие детали) и к комбинированию их в нужные динамические функциональные структуры (иначе говоря, к синтезу их в целые функциональные системы).
Таким образом, этот функциональный блок головного мозга обладает высокой модальной специфичностью: входящие в его состав части приспособлены к тому, чтобы принимать зрительную, слуховую, вестибулярную или общечувствительную информацию. В системы этого блока включаются и центральные аппараты вкусовой и обонятельной рецепции, но у человека они настолько оттесняются центральными представительствами высших экстероцептивных, дистантных анализаторов, что занимают в коре головного мозга незначительное место.
Как мы уже указывали ранее (см. гл. I), основу этого блока составляют первичные, или проекционные, зоны коры, которые характеризуются высоким развитием нейронов IV афферентного слоя, значительная часть которых обладает высочайшей специфичностью. Ранее говорилось также, что нейроны зрительных аппаратов коры реагируют только на узкоспециальные свойства зрительных раздражителей (оттенки цвета, характер линий, направление движения).
Эти высокодифференцированные нейроны строго сохраняют модальную специфичность, и в первичной затылочной коре можно найти лишь очень небольшое число клеток, которые реагируют на звук, прикосновение или вестибулярное раздражение.
102
Следует отметить, что первичные зоны отдельных областей коры, входящих в состав этого блока, включают в свой состав и некоторое количество клеток мультимодального характера, которые реагируют на несколько видов раздражителей и, по-видимому, выполняют функцию неспецифического поддержания тонуса; однако эти клетки составляют лишь небольшую часть всего нейронного состава первичных зон коры (по некоторым данным, количество их не превышает 4—5 %).
Над первичными, или проекционными, зонами коры разбираемого функционального блока мозга (составляющими его основу) надстроены, как мы видели ранее, аппараты вторичных, или гностических, зон коры, в которых IV афферентный слой уступает ведущее место II и III слоям, не имеющим столь выраженной модальной специфичности и включающим в свой состав значительное число ассоциативных нейронов с короткими аксонами, что позволяет комбинировать поступающие возбуждения в нужные «функциональные узоры» и осуществлять синтетическую функцию.
Подобное иерархическое строение, как уже отмечалось ранее, в той или иной степени свойственно всем областям коры, включенным во второй функциональный блок мозга.
В зрительной (затылочной) коре над первичными зрительными зонами (17-е поле Бродмана) надстроены вторичные зрительные поля (18-е и 19-е поля Бродмана), которые, сохраняя свою модальную (зрительную) специфичность, работают в качестве аппарата, организующего те зрительные возбуждения, которые поступают в первичные зрительные поля.
С принципами работы зрительного анализатора мы познакомимся при рассмотрении нарушений зрительного восприятия, наблюдаемых в клинике локальных поражений этих зон мозговой коры.
Слуховая (височная) кора построена по тому же принципу. Ее первичные (проекционные) зоны скрыты в глубине височной коры и поперечных извилинах Гешля (41-е поле Бродмана). Нейроны этих зон высокодифференцированны и модально специфичны. Реагируют они только на те или иные звуковые раздражители. Как и первичное зрительное поле, первичные отделы слуховой коры имеют четкое топографическое строение. Как полагает ряд авторов, волокна, несущие возбуждения от тех отделов Кортиева органа, которые реагируют на высокие тона, располагаются во внутренних (медиальных), а волокна, реагирующие на низкие тона, — в наружных (латеральных) отделах Гешлевской извилины. Отличие первичных зон слуховой коры состоит в том, что соответствующие аппараты Кортиева органа представлены в проекционных зонах слуховой коры обоих полушарий, хотя преимущественно контралатеральный характер этого представительства все же сохраняется.
103
Над аппаратами первичной слуховой коры надстроены аппараты вторичной слуховой коры, которые расположены во внешних (конвекситальных) отделах височной области (22-, 42- и частично 21-е поля Бродмана) и которые также преимущественно состоят из II и III слоев клеток. Здесь, как и в соответствующих аппаратах зрительной коры, происходит превращение соматотопических проекций импульсов в их функциональную организацию. На детальном анализе функций этих зон и на картинах нарушений слухового восприятия, которые возникают при их поражении, мы остановимся далее.
Принципиально та же функциональная организация сохраняется, наконец, и в общечувствительной (теменной) коре. Как и в случае зрительной и слуховой коры, ее основу составляет первичная, или проекционная, зона (3-е поле Бродмана), где широко представлены высокомодально-специфичные нейроны IV слоя; для этого участка коры характерна и столь же четкая соматотопическая топография, в результате чего раздражение верхних участков этой зоны вызывает появление кожных ощущений в нижних конечностях, средних участков — в верхних конечностях контралатеральной стороны, а раздражение пунктов нижнего пояса этой зоны — ощущения в контралатеральных отделах лица, губ и языка.
Над первичной зоной общечувствительной (теменной) коры надстраиваются ее вторичные зоны (1-, 2- и 5-е поля Бродмана); так же, как и вторичные зоны зрительного и слухового анализаторов, эти зоны состоят преимущественно из нейронов II и III (ассоциативных) слоев, и их раздражение приводит к возникновению комплексных кожных и кинестетических ощущений. Эффект локального поражения их будет впоследствии предметом нашего специального рассмотрения.
Таким образом, основные, модально-специфические зоны разбираемого нами второго блока мозга, каждая из которых представляет собой центральный корковый аппарат того или иного модально-специфического анализатора, построены по единому принципу иерархической организации, сформулированному в свое время еще Кэмпбеллом (1905) (рис. 41). В дальнейшем представления о цитоархитектонической структуре коры мозга человека были существенно уточнены. На рисунке 42 приводится принятая сейчас карта корковых полей, разработанная Московским институтом мозга.
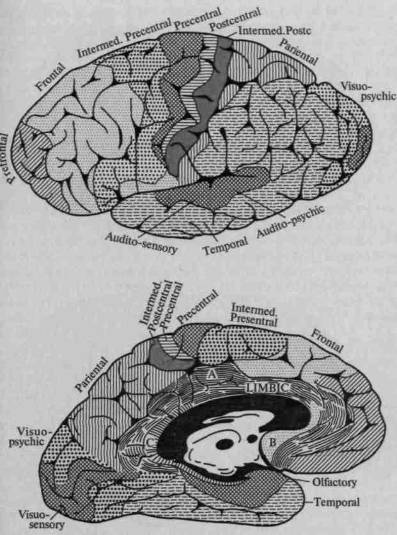
Рис. 41. Карта иерархического строения полей коры головного мозга
(по Кэмпбеллу, 1905):
вверху — наружная поверхность; внизу — внутренняя поверхность
Рис. 42. Карта цитоархитектонических полей Бродмана и Московского
института мозга: а — наружная поверхность; б — внутренняя поверхность
Как мы уже говорили ранее, познавательная деятельность человека никогда не протекает, опираясь лишь на одну изолированную модальность (зрение, слух, осязание); любое предметное восприятие (и тем более представление) является результатом полимодальной деятельности, которая первоначально в онтогенезе имеет развернутый характер и лишь затем становится свернутой. Поэтому естественно, что она должна опираться на совместную работу целой системы зон коры головного мозга.
104
Функцию обеспечения такой совместной работы целой группы анализаторов несут третичные зоны обсуждаемого нами блока, или, как их принято обозначать, зоны перекрытия корковых отделов различных анализаторов. Эти зоны расположены на границе затылочного, височного и заднецентрального отделов коры; их основную часть составляют образования нижнетеменной обла-
105
ста, которая у человека развита настолько, что составляет едва ли не четвертую часть всех образований описываемого блока. Это дает основание считать третичные зоны («задний ассоциативный центр», как их обозначал Флексиг) специфически человеческими образованиями.
106
Как уже говорилось, третичные зоны задних отделов мозга состоят почти целиком из клеток II и III (ассоциативных) слоев коры и, следовательно, функция их почти полностью сводится к интеграции возбуждений, приходящих из разных анализаторов. Есть основания думать, что подавляющая часть нейронов этих зон имеет мультимодальный характер и реагирует на комплексные признаки среды (например, на признаки пространственного расположения, количество элементов), на которые не реагируют нейроны первичных и даже вторичных зон коры. Именно к таким третичным образованиям задних отделов мозговой коры относятся 7-, 39- и 40-е поля Бродмана (верхние и нижние отделы теменной области), 21-е поле височной и 37-е поле височно-затылочной областей.
Далее мы рассмотрим функции третичных образований описываемых отделов коры подробнее и убедимся на основании анализа психологических экспериментов и клинических данных, что в основном эти зоны связаны с пространственной организацией возбуждений и с превращением последовательно поступающих (сукцессивных) сигналов в одновременно действующие (симультанные) группы, чем и обеспечивается тот синтетический характер восприятия, о котором в свое время упоминал И. М. Сеченов (А.Р.Лурия, 1963, 1966).
Деятельность третичных зон задних отделов коры необходима не только для успешного синтеза наглядной информации, но и для перехода от уровня непосредственного наглядного синтеза к уровню символических процессов, для оперирования значениями слов, сложными грамматическими и логическими структурами, системами чисел и отвлеченными соотношениями. Другими словами, третичные зоны задних отделов коры являются аппаратами, участие которых необходимо для превращения наглядного восприятия в отвлеченное мышление, опосредствованное всегда внутренними схемами, и для сохранения в памяти организованного опыта.
Совокупность данных позволяет назвать этот функциональный блок мозга блоком получения, переработки и хранения экстероцептивной информации. Можно указать некоторые законы построения коры, входящей в состав второго блока мозга (этими законами описывается также и строение третьего блока).
Первый закон — закон иерархического строения корковых зон. Соотношение первичных, вторичных и третичных зон коры, осуществляющих все более сложный синтез приходящей информации, является достаточно отчетливой иллюстрацией этого закона.
Следует отметить, что отношения между первичными, вторичными и третичными зонами коры, входящими в состав этого блока, не остаются одинаковыми в процессе онтогенетического развития. Так, у маленького р е б е н к а для успешного фор-
107
мирования вторичных зон необходима сохранность первичных зон, а для формирования третичных зон — достаточная сформированность вторичных зон коры. Поэтому нарушение в раннем возрасте низших зон коры соответствующих типов неизбежно приводит к недоразвитию более высоких зон коры; таким образом, как это было сформулировано Л. С. Выготским (1934, 1960), основная линия взаимодействия этих зон коры направлена «снизу вверх».
Наоборот, у взрослого человека с полностью сложившимися психологическими функциями ведущее место переходит к высшим зонам коры. Даже воспринимая окружающий мир, взрослый человек организует свои впечатления в логические системы; иными словами, наиболее высокие, третичные, зоны коры у него управляют работой подчиненных им вторичных зон, а при поражении последних оказывают на их работу компенсирующее влияние. Такой характер взаимоотношений иерархически построенных зон коры в зрелом возрасте позволил Л. С. Выготскому заключить, что на позднем этапе онтогенеза они взаимодействуют «сверху вниз».
Второй закон можно сформулировать как закон убывающей специфичности иерархически построенных зон коры. Как уже было показано, первичные зоны коры каждой из частей, входящих в состав описываемого блока, обладают максимальной модальной специфичностью. Это характерно для первичных (проекционных) зон как зрительной, так и слуховой и общечувствительной коры. Основа этого явления — наличие огромного числа нейронов с высокодифференцированной, модально-специфической функцией.
Вторичные зоны коры, где преобладают верхние слои с их ассоциативными нейронами, обладают модальной специфичностью в значительно меньшей степени.
Будучи тесно связанными с корковыми отделами соответствующих анализаторов, эти зоны (которые Г. И. Поляков (1969) предпочитает называть проекционно-ассоциативными) характеризуются модально-специфическими гностическими функциями. Здесь интегрируется в одних случаях зрительная (вторичные затылочные зоны), в других — слуховая (вторичные височные зоны), в третьих — тактильная информация (вторичные теменные зоны). Однако тот факт, что эти зоны (в которых, как уже говорилось, преобладают мультимодальные нейроны и нейроны с короткими аксонами) играют основную роль в превращении соматотипической проекции в функциональную организацию поступающей информации, указывает на то, что степень специализированности клеток этих зон значительно меньше, чем в первичных зонах, и что переход к ним знаменует существенный шаг в направлении «убывания модальной специфичности».
108
Еще в меньшей степени модальная специфичность характеризует третичные зоны описываемого блока, которые можно обозначить как «зоны перекрытия» корковых отделов различных анализаторов. Модальная специфичность входящих в их состав элементов представлена еще меньше, симультанные (пространственные) синтезы, которые осуществляются этими зонами коры, трудно расценить как модально-специфические. В еще меньшей степени этот принцип можно относить к высшим, символическим уровням работы этих зон, с переходом к которым функция третичных зон в известной мере приобретает надмодальный характер.
Таким образом, закон убывающей специфичности является другой стороной упомянутого нами ранее закона иерархического строения входящих в состав второго блока зон коры, обеспечивающих переход от дробного отражения частных модально-специфических признаков к синтетическому отражению более общих и отвлеченных схем воспринимаемого мира.
Следует отметить, что вторичные и третичные зоны коры (в которых преобладают мультимодальные и ассоциативные нейроны и которые не имеют прямой связи с периферией) имеют не менее важное функциональное значение, чем первичные зоны. Несмотря на убывающую специфичность (а может быть, как раз в силу такой убывающей специфичности), они приобретают способность играть организующую, интегрирующую роль в работе более специфических зон, становятся ответственными за организацию функциональных систем, необходимых для осуществления сложных познавательных процессов.
Без учета этого все клинические факты функциональных нарушений, возникающих при локальных поражениях мозга (которые будут описаны далее), останутся для нас непонятными.
Нам остается сформулировать третий закон организации описываемого нами функционального блока, который оказывается, впрочем, приложимым и к организации коры головного мозга в целом. Его можно обозначить как закон прогрессивной латерализации функций, т.е. связи функций с определенным полушарием мозга по мере перехода от первичных зон коры к вторичным и затем третичным зонам.
Известно, что первичные зоны обоих полушарий мозга равноценны: и те и другие являются проекциями контралатеральных (расположенных на противоположной стороне) воспринимающих поверхностей, и ни о каком доминировании первичных зон того или другого полушария говорить нельзя.
Иначе обстоит дело при переходе к вторичным, а затем третичным зонам. Известно, что с возникновением праворукости (а ее появление связано с трудом и, по-видимому, относится к очень ранним этапам истории человека), а затем и связанной с ней речи, возникает известная латерализация функций, которая отсутствует у животных, но которая у человека становится важным принципом функциональной организации мозга.
109
Левое полушарие (у правшей) становится доминантным; именно оно начинает осуществлять речевые функции, в то время как правое полушарие, не связанное с деятельностью правой руки и речью, остается субдоминантным. Этот принцип латерализации функций становится решающим принципом функциональной организации мозговой коры человека.
Левое (доминантное) полушарие играет существенную роль не только в мозговой организации самих речевых процессов, но и в мозговой организации всех связанных с речью высших форм психической деятельности — категориального восприятия, активной речевой памяти, логического мышления и др., в то время как правое (субдоминантное) полушарие в меньшей степени участвует в их протекании.
Принцип латерализации высших функций в коре головного мозга вступает в силу только на уровне вторичных и третичных зон коры, которые играют основную роль в функциональной организации информации, доходящей до коры головного мозга, осуществляемой у человека с помощью речи. У взрослого человека функции вторичных и третичных зон левого (ведущего) полушария начинают коренным образом отличаться от функций вторичных и третичных зон правого (субдоминантного) полушария. Подавляющее число симптомов нарушения высших психологических процессов, описанных при локальных поражениях мозга, относится к симптомам, возникающим при поражениях вторичных и третичных зон доминантного (левого) полушария, в то время как симптоматика поражения этих же зон субдоминантного (правого) полушария изучена несравненно меньше. Проблема своеобразия в организации функций мозга человека снова привлекает к себе острое внимание исследователей (см. обзор Дрю, Эттлингера, Милнер и Пассингхэма, 1970).
Следует, однако, учитывать, что абсолютная доминантность левого полушария встречается далеко не всегда и, таким образом, закон латерализации имеет лишь относительный характер. Так, по данным последних исследований (Зангвилл, 1960; Субирана, 1969), лишь одна четвертая часть людей является полностью правшами, несколько больше одной трети проявляет выраженное преобладание левого полушария, в то время как остальные отличаются относительно слабым преобладанием левого полушария, а в одной десятой всех случаев такое преобладание левого полушария вообще отсутствует.
Разбирая далее основные картины изменений высших психических процессов при поражениях отдельных зон мозговой коры, мы еще будем иметь случай наглядно убедиться в этом положении.
110
Резюмируем сказанное выше.
Вторым функциональным блоком коры головного мозга является блок приема, переработки и хранения экстероцептивной информации. Он расположен в задних отделах полушарий и включает в свой состав зрительные (затылочные), слуховые (височные) и общечувствительные (теменные) отделы коры головного мозга и соответствующие подкорковые структуры.
Аппараты этого (как и следующего) блока имеют иерархическое строение, распадаясь на первичные (проекционные) зоны, которые принимают информацию и дробят ее на мельчайшие составные части, вторичные (проекционно-ассоциативные) зоны, которые обеспечивают кодирование (синтез) этих составных частей и превращают соматотопическую проекцию в функциональную организацию, и третичные зоны (или зоны перекрытия), обеспечивающие совместную работу различных анализаторов и выработку надмодальных (символических) схем, лежащих в основе комплексных форм познавательной деятельности.
Указанные иерархически построенные зоны коры разбираемого блока работают по принципам убывающей модальной специфичности и возрастающей функциональной латерализации. Оба эти принципа и обеспечивают возможность наиболее сложных форм работы мозга, лежащих в основе наиболее высоких видов познавательной деятельности человека, генетически связанных с трудом, а структурно — с участием речи в организации психических процессов.
Таковы самые общие принципы работы второго функционального блока головного мозга.
3 БЛОК ПРОГРАММИРОВАНИЯ, РЕГУЛЯЦИИ И КОНТРОЛЯ СЛОЖНЫХ ФОРМ ДЕЯТЕЛЬНОСТИ
Прием, переработка и хранение внешней информации составляют только одну сторону психической жизни человека. Ее другую сторону составляет организация активной сознательной психической деятельности. С этой задачей и связан третий из основных функциональных блоков мозга — блок программирования, регуляции и контроля за протекающей деятельностью.
Человек не только пассивно реагирует на поступающие сигналы. Он формирует планы и программы своих действий, следит за их выполнением и регулирует свое поведение, приводя его в соответствие с этими планами и программами; наконец, он контролирует свою сознательную деятельность, сличая эффект своих действий с исходными намерениями и корригируя допущенные им ошибки.
111
Все эти процессы активной сознательной деятельности требуют совсем иных мозговых аппаратов, чем те, которые мы описали ранее. Если даже в простых рефлекторных актах наряду с афферентной стороной существует эффекторная сторона и аппараты обратной связи служат контрольным сервомеханизмом, то тем более необходимы такие специальные контролирующие нервные образования в сложных психических актах. Этим задачам и служат аппараты третьего блока головного мозга, функцию которого мы уже обозначили выше.
Аппараты третьего функционального блока расположены в передних отделах больших полушарий, спереди от передней центральной извилины (рис. 43).
Рис. 43. Схема расположения лобных
(префронтальных) отделов мозга
у человека
«Выходными воротами» этого блока является двигательная зона коры (4-е поле Бродмана), V слой которой содержит гигантские пирамидные клетки Беца, волокна от которых идут к двигательным ядрам спинного мозга, а оттуда к мышцам, составляя часть большого пирамидного пути. Эта зона коры, как мы уже видели (рис. 23), топографически построена так, что ее верхние отделы являются источником волокон, идущих к нижним конечностям противоположной стороны, средние — к верхним конечностям противоположной стороны, а нижние — волокон, направляющихся к мышцам лица, губ и языка. Мы уже говорили, что в значительной степени в этой зоне представлены органы, имеющие наибольшее функциональное значение и нуждающиеся в наиболее тонкой регуляции.
Первичная двигательная кора не может работать изолированно; все движения человека в той или иной степени нуждаются в тоническом пластическом фоне, который обеспечивается базаль-ными двигательными узлами и волокнами экстрапирамидной системы. Значение этой системы, обеспечивающей фон всех произвольных движений, служило предметом многочисленных исследований, и мы не будем останавливаться на них особо.
Первичная двигательная кора является, как мы сказали, «выходными воротами» двигательных импульсов, или, как говорил выдающийся исследователь движений Н. А. Бернштейн, «передними рогами головного мозга». Естественно, однако, что двигательный состав тех импульсов, которые она посылает на периферию, должен быть хорошо подготовлен, включен в определенные программы. Без такой подготовки импульсы, направляемые через переднюю центральную извилину, не могут обеспечить целесообразные движения.
Подготовка двигательных импульсов не может быть выполнена самими пирамидными клетками: она должна быть обеспечена как в аппарате самой передней центральной извилины, так и в аппаратах надстроенных над ней вторичных зон двигательной коры, которые готовят двигательные программы, лишь затем передающиеся на гигантские пирамидные клетки.
112
В пределах передней центральной извилины аппаратом, участвующим в подготовке двигательных программ и передаче их на гигантские пирамидные клетки, являются верхние слои коры и гли-альное серое вещество, окружающее нейроны. Как было в свое время показано Бенином (1943, 1948), отношение массы глиального вещества к массе нейронов передней центральной извилины резко возрастает по мере эволюции, так что величина глиального коэффициента у человека вдвое больше, чем у высших обезьян, и почти в пять раз больше, чем у низших (табл. 8).
Таблица 8
Дата: 2018-11-18, просмотров: 1168.