Физическая модель биоткани
Четыре независимых макроскопических параметра характеризуют распространение излучения в ткани: параметр анизотропии рассеяния (g), коэффициент поглощения (ma), коэффициент рассеяния (ms), показатель преломления (n) (см. табл. 1). Перечисленные оптические параметры содержат информацию как о биохимических свойствах, так и морфологических, структурных и функциональных особенностях ткани [1,3,4].
Таблица 1
Макроскопические оптические параметры, характеризующие распространение излучения в биоткани
| Параметр | Обозначение | Характеристика |
| Показатель преломления | n | Отношение скорости излучения в вакууме к скорости в среде |
| Коэффициент поглощения (см-1) | mа | Величина, обратная средней длине свободного пробега поглощенного фотона в биоткани |
| Коэффициент рассеяния (см-1) | ms | Величина, обратная средней длине свободного пробега однократно рассеянного фотона в биоткани |
| Фактор анизотропии | g | Средний косинус угла рассеяния |
| Транспортный коэффициент рассеяния (см-1) | ms’ ms’=ms(1-g) | Величина, обратная средней длине изотропного рассеяния |
Математически параметр анизотропии рассеяния определяется как средний косинус угла рассеяния q
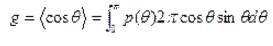
где p(q) = p(s,s') - фазовая функция; p(s,s') описывает рассеивающие свойства среды и представляет собой функцию плотности вероятности для рассеяния в направлении s'фотона, движущегося в направлении s, т.е. характеризует элементарный акт рассеяния. Если рассеяние симметрично относительно направления падающей волны, тогда фазовая функция зависит только от угла q между направлениями s и s'.
Предположение о случайном распределении рассеивателей в среде, что означает отсутствие в структуре биоткани пространственной корреляции, ведет к следующей нормировке
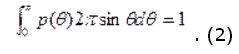
Во многих практических случаях фазовая функция хорошо аппроксимируется с помощью функции Хеньи-Гринштейна:
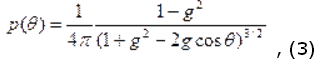
Значение g изменяется в пределах от 0 до 1: g = 0 соответствует случаю изотропного (рэлеевского) рассеяния, g = 1 – полному рассеянию вперед (рассеяние Ми на крупных частицах). Для биологических тканей g = 0,7-0,99.
Величина, обратная средней длине свободного пробега (СДСП) является удобным способом описания коэффициентов рассеяния и поглощения. СДСП однократно рассеянного фотона в биоткани определяется как lph = mt-1 = (ma+ ms)-1; mt - коэффициент экстинкции (коэффициент ослабленияизлучения).
Транспортный (редуцированный) коэффициент рассеяния (ms') не является независимым от других параметров в табл. 1 и определяется как
ms' = ms × (1- g). (4)
Транспортный коэффициент рассеяния определяет характерную длину, на которой фотоны теряют память о своем первоначальном направлении, т.е. ms' дает информацию об изотропно-рассеянных событиях.
Средняя транспортная длина пробега фотона 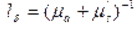
Отметим, что средняя транспортная длина пробега фотона в среде с анизотропным однократным рассеянием существенно выше длины свободного пробега в среде с изотропным однократным рассеянием 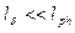
Транспортная длина 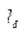 означает такую длину, на которой фотон теряет свое первоначальное направление.
означает такую длину, на которой фотон теряет свое первоначальное направление.
Механизмы передачи энергии
Энергия g-квантов, испускание которых сопровождает ядерные превращения подавляющего большинства радионуклидов, определяется дискретными уровнями возбужденных ядер и составляет, как правило, от ~ 10 кэВ до ~ 6 МэВ. Для электромагнитных квантов (фотонов), имеющих такую энергию, основными первичными процессами взаимодействия с веществом являются фотопоглощение, комптоновское рассеяние и образование пар электрон-позитрон (рис. 3.16). При этом часть энергии преобразуется в энергию заряженных частиц, часть – во вторичное фотонное излучение.
Фотопоглощение (фотоэффект) – процесс передачи всей энергии g-кванта электрону, как правило, К-оболочки атома облучаемого вещества. При этом электрон (фотоэлектрон) выбрасывается из атома, его кинетическая энергия (Еф) равна разности между энергией фотона и энергией связи в атоме (EK): Еф = Eγ−EK.
Рассмотрение законов сохранения энергии и импульса показывает, что фотоэффект невозможен на свободном электроне. Вероятность фотоэффекта существенно возрастает по мере увеличения энергии связи электрона в атоме. Когда энергия фотона достаточна для ионизации К-оболочки (Eγ > EK), то вероятность фотоэффекта на К-оболочке выше, чем на L-оболочке и M-оболочке примерно в 5 и в 20 раз, соответственно. Очевидно, что при Eγ< EK фотоэффект возможен только на L-, M-,… и т.д. оболочках.
Вакансия, образовавшаяся на К-оболочке, заполняется электроном с одного из находящихся выше уровней, например, LI (рис. 3.17). Выделяющаяся при этом энергия (EK−ELI) может быть либо рассеяна в виде рентгеновского кванта (EX = EK−ELI), либо передана другому электрону, например, LII-оболочки. Этот электрон покинет атом, т.к. EK−ELI>ELII. В этом заключается эффект Оже. Каскадное размножение «дырок» после первого оже-перехода происходит до тех пор, пока они не переместятся во внешние оболочки. Таким образом, фотоэффект сопровождается либо испусканием характеристических рентгеновских квантов, либо каскадом безизлучательных переходов. Вероятность испускания оже-электронов увеличивается с уменьшением Z, и для легких атомов (Z<30) она выше вероятности рентгеновской флуоресценции.
Комптоновское рассеяние– передача части энергии g-квантов электронам внешних оболочек. Энергия связи этих электронов пренебрежимо мала по сравнению с Eγ и рассеяние трактуется как столкновение фотона с отдельным свободным электроном. В результате электрон покидает атом, а новый γ-квант с энергией hν′=Eγ′ отклоняется от направления движения первичного кванта (рис. 3.13б, 0º<θ£180º). Энергия рассеянного кванта изменяется в соответствии с (3.21)
Классическое или томсоновское рассеяние. Эффект Комптона не существенен для мягкого (длинноволнового) излучения. Однако следует отметить, что γ-излучение (как и рентгеновское) может рассеиваться и без изменения длины волны: фотон поглощается и немедленного испускается атомом без изменения энергии, но в другом направлении. Рассеяние излучения без передачи энергии облучаемому веществу называют классическим или томсоновским. Оно возможно, когда энергия фотона недостаточна для вырывания электрона из атома, и впервые наблюдалось для мягкого рентгеновского излучения (λ≈10−8 см, Е ≤ 10 кэВ). Его источником являются связанные электроны атома, которые приходят в резонансные колебания под действием падающего излучения и вследствие этого сами становятся излучателями квантов такой же частоты.
Образование пар электрон-позитрон– взаимодействиевысокоэнергетических g-квантов с полем ядер, приводящее к исчезновению квантов и образованию заряженных частиц. Пороговая энергия процесса равна удвоенной энергии массы покоя электрона (1022 кэВ), а кинетическая энергия частиц Ee- = Ee+= (Eγ-1022)/2. Последующая аннигиляция позитрона приводит к появлению двух квантов с энергией по 511 кэВ каждый, которые, в свою очередь, могут поглотиться веществом по механизму фотоэффекта или рассеяться на электронах. Сечение образования пар (c) пропорционально Z2 и растет с увеличением Eγ. В интервале 2550 <Eγ <25000 кэВ значение cµ Z2∙lnEγ
Фотоизомеризация молекулы
Химические свойства молекулы определяются не только составом входящих в нее атомов, но и их взаимным расположением. Молекулы с одинаковым составом и разной пространственной конфигурацией называются изомерами. Их химические свойства различны.
Фотоизомеризация - изменение пространственной структуры молекулы, возникающее после ее фотовозбуждения.
Упрощенная схема фотоизомеризации показана на рис. Молекула поглощает фотон и переходит в возбужденное состояние с сохранением первоначальной конфигурации. Взаимодействие с окружающими молекулами приводит к безызлучательному снятию части возбуждения с одновременным изменением пространственной конфигурации. Заканчивается процесс образованием изомера в основном (невозбужденном) состоянии.
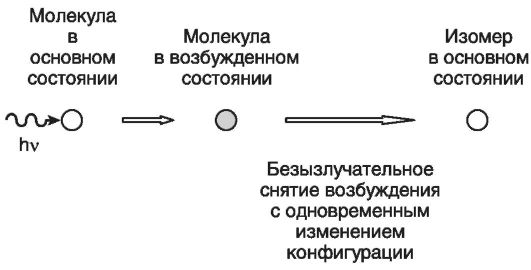
Молекула поглощает фотон и переходит в возбужденное состояние с сохранением первоначальной конфигурации. Взаимодействие с окружающими молекулами приводит к безизлучательному снятию части возбуждения с одновременным изменением пространственной конфигурации. Заканчивается процесс образованием изомера в основном (невозбужденном) состоянии.
Примером пространственных изомеров являются цис- и трансизомеры, в молекулах которых заместители у атомов углерода расположены соответственно по одну и по разные стороны плоскости двойной связи (рис. 30.2).
Переход от одного изомера к другому связан с поворотом на 180° вокруг двойной связи.
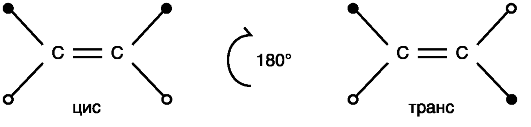
Рис. Цис- и транс-изомеры
У человека цис-транс-фотоизомеризации пигмента ретиналя принадлежит основная роль в зрительной рецепции.
Оптические свойства биотканей.
Ткани - необычайно сложные комплексы, включающие огромное разнообразие молекул, структур и функциональных единиц. Несмотря на такую сложность, возможно рассмотрение усредненных оптических свойств с учетом определенных ограничений и допущений.
С оптической точки зрения биоткани, включая биологические жидкости: кровь, лимфу и пр., - можно разделить на два больших класса:
1) сильно рассеивающие (оптически мутные), такие как кожа, мозг, стенка сосуда, кровь, склера, оптические свойства которых описываются моделью многократного рассеяния;
2) слабо рассеивающие (прозрачные), такие, как роговица и хрусталик глаза, оптические свойства которых описываются в модели однократного (или малократного) рассеяния [1].
Биологические ткани являются оптически неоднородными сильно рассеивающими и поглощающими средами со средним показателем преломления, большим, чем у воздуха. На границе раздела биообъект - воздух часть излучения отражается, так называемое френелевское отражение, а остальная часть проникает в биоткань. За счет многократного рассеяния и поглощения излучение экспоненциально затухает при распространении в биоткани. Объёмное рассеяние является причиной распространения значительной доли излучения в обратном направлении, обратное рассеяние.
Поглощенный свет преобразуется в тепло, переизлучается в виде флуоресценции, а также тратится на фотобиохимические реакции. Спектр поглощения определяется типом доминирующих поглощающих центров, а также содержанием воды в биоткани. В ультрафиолетовой (УФ, λ<390 нм) и инфракрасной (ИК) (λ>1 мкм) областях спектра велико поглощение белками и водой, соответственно, поэтому вклад рассеяния сравнительно мал, и свет неглубоко проникает в биоткань – всего на несколько клеточных слоев [4]. Для коротких волн видимой области глубина проникновения типичной биоткани составляет 0,5 ¸ 2,5 мм; имеет место как поглощение, так и рассеяние; около 15 ¸ 40% падающего излучения отражается. В области длин волн 0,6¸0,9 мкм рассеяние превалирует над поглощением, следовательно, глубина проникновения света увеличивается до 8¸20 мм. Также существенно увеличивается интенсивность отраженного и рассеянного в обратном направлении биотканью излучения, до 35-70% от падающего [1,4]. В зеленой области спектра 500-570 нм доминирует поглощение гемоглобном крови.
Из-за многослойной и многокомпонентной структуры кожи взаимодействие света с ней оказывается весьма сложным. Роговой слой отражает около 5 ¸ 7% падающего излучения. Коллимированный пучок света преобразуется в диффузный за счет микроскопических неоднородностей на границе воздух – роговой слой. Большая часть отраженного кожей света образуется за счет обратного рассеяния различными слоями ткани (роговой слой, эпидермис, дерма и микрососудистая система). Поглощение рассеянного света пигментами кожи дает количественную информацию о концентрации билирубина, меланина, воды, о насыщении гемоглобина кислородом, о содержании лекарственных препаратов и других поглотителей в ткани и крови, что является основой ряда диагностических методов. Значительное проникновение видимого и ближнего ИК света через кожу внутрь организма человека, в област Твердые ткани, такие, как ребра и черепная коробка, а также цельная кровь демонстрируют сравнительно хорошее пропускание в видимой и ближней ИК области спектра [1,4]. и длин волн так называемого “терапевтического окна” (650 ¸ 900 нм), является основой ряда методов фототерапии и оптической диагностики (спектрофотометрии).
Основные биологические хромофоры
Хромофоры биологических тканей, вещества, ответственные за поглощение излучения, могут быть разделены на те, которые проявляют кислородозависимое поглощение (окси- и дезоксигемоглобин, миоглобин и цитохромоксидаза), и те, чьё поглощение существенно не меняется в процессе клинических измерений in vivo (вода, меланин, билирубин, жировая ткань) [5,6].
Анализ литературных данных [5,6] и расчеты по соответствующим концентрациям и молярным коэффициентам экстинкции позволяют установить, что основной вклад в общий коэффициент поглощения в красной и ближней ИК областях спектра вносят фракции гемоглобина и вода. Вклад остальных хромофоров колеблется от долей процента для билирубина и до 10% для цитохромоксидазы и меланина [6]. Корректность учета вклада этих хромофоров в общий коэффициент поглощения определяет составляющую методической погрешности спектрофотометерических методов, что по-разному учитывается в различных методах спектрофотометрии и их приборных реализациях.
Физическая модель биоткани
Четыре независимых макроскопических параметра характеризуют распространение излучения в ткани: параметр анизотропии рассеяния (g), коэффициент поглощения (ma), коэффициент рассеяния (ms), показатель преломления (n) (см. табл. 1). Перечисленные оптические параметры содержат информацию как о биохимических свойствах, так и морфологических, структурных и функциональных особенностях ткани [1,3,4].
Таблица 1
Макроскопические оптические параметры, характеризующие распространение излучения в биоткани
| Параметр | Обозначение | Характеристика |
| Показатель преломления | n | Отношение скорости излучения в вакууме к скорости в среде |
| Коэффициент поглощения (см-1) | mа | Величина, обратная средней длине свободного пробега поглощенного фотона в биоткани |
| Коэффициент рассеяния (см-1) | ms | Величина, обратная средней длине свободного пробега однократно рассеянного фотона в биоткани |
| Фактор анизотропии | g | Средний косинус угла рассеяния |
| Транспортный коэффициент рассеяния (см-1) | ms’ ms’=ms(1-g) | Величина, обратная средней длине изотропного рассеяния |
Математически параметр анизотропии рассеяния определяется как средний косинус угла рассеяния q
где p(q) = p(s,s') - фазовая функция; p(s,s') описывает рассеивающие свойства среды и представляет собой функцию плотности вероятности для рассеяния в направлении s'фотона, движущегося в направлении s, т.е. характеризует элементарный акт рассеяния. Если рассеяние симметрично относительно направления падающей волны, тогда фазовая функция зависит только от угла q между направлениями s и s'.
Предположение о случайном распределении рассеивателей в среде, что означает отсутствие в структуре биоткани пространственной корреляции, ведет к следующей нормировке
Во многих практических случаях фазовая функция хорошо аппроксимируется с помощью функции Хеньи-Гринштейна:
Значение g изменяется в пределах от 0 до 1: g = 0 соответствует случаю изотропного (рэлеевского) рассеяния, g = 1 – полному рассеянию вперед (рассеяние Ми на крупных частицах). Для биологических тканей g = 0,7-0,99.
Величина, обратная средней длине свободного пробега (СДСП) является удобным способом описания коэффициентов рассеяния и поглощения. СДСП однократно рассеянного фотона в биоткани определяется как lph = mt-1 = (ma+ ms)-1; mt - коэффициент экстинкции (коэффициент ослабленияизлучения).
Транспортный (редуцированный) коэффициент рассеяния (ms') не является независимым от других параметров в табл. 1 и определяется как
ms' = ms × (1- g). (4)
Транспортный коэффициент рассеяния определяет характерную длину, на которой фотоны теряют память о своем первоначальном направлении, т.е. ms' дает информацию об изотропно-рассеянных событиях.
Средняя транспортная длина пробега фотона
Отметим, что средняя транспортная длина пробега фотона в среде с анизотропным однократным рассеянием существенно выше длины свободного пробега в среде с изотропным однократным рассеянием
Транспортная длина означает такую длину, на которой фотон теряет свое первоначальное направление.
Дата: 2018-12-21, просмотров: 968.