Глюконеогенез – процесс синтеза глюкозы из веществ неуглеводной природы.
Основная функция глюконеогенеза – поддержание постоянного уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок.
Глюконеогенез включает все механизмы и пути, обеспечивающие образование глюкозы и гликогена из неуглеводных компонентов. Главными субстратами глюконеогенеза служат глюкогенные аминокислоты, лактат, глицерол и (у жвачных) пропионат. Гликонеогенез происходит главным образом в печени и почках, поскольку именно в этих органах имеется полный набор необходимых ферментов.
Глюконеогенез обеспечивает потребности организма в глюкозе в тех случаях, когда диета содержит недостаточное количество углеводов. Постоянное поступление глюкозы в качестве источника энергии особенно необходимо для нервной системы и эритроцитов. При понижении концентрации глюкозы в крови ниже определенного критического уровня нарушается функционирование мозга; при тяжелой гипогликемии возникает коматозное состояние и может наступить летальный исход. Глюкоза необходима также для жировой ткани как источник глицерола, входящего в состав глицеридов; она играет, вероятно, существенную роль в поддержании эффективных концентраций интермедиатов цикла лимонной кислоты во многих тканях. Из этого следует, что даже в условиях, когда большая часть потребностей организма в калориях обеспечивается за счет жира, всегда сохраняется определенная потребность в глюкозе. Кроме того, глюкоза служит единственным видом топлива для работы скелетной мышцы в анаэробных условиях. Она является предшественником молочного сахара (лактозы) в молочных железах и активно потребляется плодом в период развития. Следует отметить также, что механизм глюконеогенеза используется для удаления из крови продуктов тканевого метаболизма, например лактата, образующегося в мышцах и эритроцитах, глицерола, непрерывно образующегося в жировой ткани.
Пропионат – главная глюкогенная жирная кислота, образующаяся в процессе переваривания углеводов жвачными животными, является главным субстратом глюконеогенеза у этих животных.
Метаболические пути, участвующие в глюконеогенезе, являются модификациями путей гликолиза и цикла лимонной кислоты. Кребс отметил, что простому обращению гликолиза препятствуют энергетические барьеры на ряде стадий:
1) между пируватом и фосфоенолпируватом,
2) между фруктозо-1,6-бисфосфатом и фруктозо-6-фосфатом,
3) между глюкозо-6-фосфатом и глюкозой,
4) между глюкозо-1 -фосфатом и гликогеном.
Эти барьеры обходятся с помощью специальных реакций.
1) В митохондриях имеется фермент пируваткарбоксилаза, который при участии АТР, биотина (витамина группы В) и СО2 превращает пируват в оксалоацетат. Функция биотина заключается в присоединении СО2 (из бикарбоната) к ферменту, далее СО2 переносится на пируват. Во внемитохондриальной среде клетки имеется второй фермент – фосфоенолпнруваткарбоксикиназа, который катализирует превращение оксалоацетата в фосфоенолпируват. Для этой реакции требуется высокоэнергетический фосфат в форме GTP или ITP; в результате реакции освобождается СО2. Таким образом, с помощью этих двух ферментов и лактатдегидрогеназы лактат может превращаться в фосфоенолпируват.
Существенное препятствие, однако, заключается в том, что выход оксалоацетата из митохондрии весьма затруднен. Оно преодолевается следующим образом: оксалоацетат превращается в соединение, легко диффундирующее из митохондрии во внемитохондриальный компартмент клетки, где это соединение снова превращается в оксалоацетат. Таким соединением служит малат; его образование из оксалоацетата внутри митохондрий и превращение обратно в оксалоацетат вне митохондрий катализируются малатдегидрогеназой.
2) Превращение фруктозо-1,6-бисфосфата во фруктозо-6-фосфат, необходимое для обращения гликолиза на рассматриваемой стадии, катализируется специфическим ферментом фруктозо-1,6-бисфосфатазой. Это – ключевой фермент в том смысле, что именно его присутствием определяется, способна ли ткань ресинтезировать гликоген из пирувата и триозофосфатов. Этот фермент имеется в печени и почках, он был также обнаружен в поперечнополосатых мышцах. Считают, что в сердечной мышце и гладких мышцах он отсутствует.
3) Превращение глюкозо-6-фосфата в глюкозу катализируется другой специфической фосфатазой – глюкозо-6-фосфатазой. Она присутствует в печени и почках, но отсутствует в мышцах и жировой ткани. Наличие этого фермента позволяет ткани поставлять глюкозу в кровь.
4) Распад гликогена с образованием глюкозо-1-фосфата осуществляется фосфорилазой. Синтез гликогена идет по совершенно другому пути, через образование уридиндифосфатглюкозы, и катализируется гликогенсинтазой.
Переключение печени с гликолиза на глюконеогенез и обратно происходит с участием инсулина и глюкагона и осуществляется с помощью:
- аллостерической регуляции активности ферментов;
- ковалентной модификации ферментов путём фосфорилирования/дефосфорилирования;
- индукции/репрессии синтеза ключевых ферментов.
«Субстратные» циклы – парные комбинации процессов синтеза и распада метаболитов.
Направление реакций первого субстратного цикла регулируется главным образом концентрацией глюкозы.
Направление реакций второго субстратного цикла зависит от активности фосфофруктокиназы и фосфатазы фруктозо-1,6-бисфосфата. Активность этих ферментов зависит от концентрации фруктозо-2,6-бисфосфата. Фруктозо-2,6-бисфосфат – метабоилит, образующийся в незначительных количествах из фруктозо-6-фосфата и выполняющий только регуляторные функции. Образование фруктозо-2,6-бисфосфата путём фосфорилирования фруктозо-6-фосфата катализирует бифункциональный фермент (БИФ), который катализирует и обратную реакцию:
 Киназная активность БИФ проявляется, когда фермент находится в дефосфорилированной форме (БИФ-ОН). Дефосфорилированная форма БИФ характерна для абсорбтивного периода, когда инсулин/глюкагоновый индекс высокий. В этот период количество фруктозо-2,6-бисфосфата увеличивается:
Киназная активность БИФ проявляется, когда фермент находится в дефосфорилированной форме (БИФ-ОН). Дефосфорилированная форма БИФ характерна для абсорбтивного периода, когда инсулин/глюкагоновый индекс высокий. В этот период количество фруктозо-2,6-бисфосфата увеличивается:
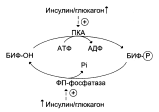 При низком инсулин/глюкагоновом индексе, характерном для периода длительного голодания, происходит фосфорилирование БИФ, и он функционирует как фосфатаза. Результат – снижение количества фруктозо-2,6-бисфосфата.
Регуляторное значение фруктозо-2,6-бисфосфата заключается в том, что он аллостерически активирует фосфофруктокиназу (фермент гликолиза). При этом фруктозо-2,6-бисфосфат снижает ингибирующее действие АТФ на этот фермент в абсорбтивном периоде и повышает его сродство к фруктозо-6-фосфату. В то же время фруктозо-2,6-бисфосфат ингибирует фруктозо-1,6-бисфосфатазу (фермент глюконеогенеза). Таким образом, в абсорбтивном периоде благодаря фруктозо-2,6-бисфосфату ускоряется процесс гликолиза.
Результатом уменьшения количества фруктозо-2,6-бисфосфата в постабсорбтивном периоде будет снижение активности фосфофруктокиназы, замедление гликолиза и переключение гликолиза на глюконеогенез.
При низком инсулин/глюкагоновом индексе, характерном для периода длительного голодания, происходит фосфорилирование БИФ, и он функционирует как фосфатаза. Результат – снижение количества фруктозо-2,6-бисфосфата.
Регуляторное значение фруктозо-2,6-бисфосфата заключается в том, что он аллостерически активирует фосфофруктокиназу (фермент гликолиза). При этом фруктозо-2,6-бисфосфат снижает ингибирующее действие АТФ на этот фермент в абсорбтивном периоде и повышает его сродство к фруктозо-6-фосфату. В то же время фруктозо-2,6-бисфосфат ингибирует фруктозо-1,6-бисфосфатазу (фермент глюконеогенеза). Таким образом, в абсорбтивном периоде благодаря фруктозо-2,6-бисфосфату ускоряется процесс гликолиза.
Результатом уменьшения количества фруктозо-2,6-бисфосфата в постабсорбтивном периоде будет снижение активности фосфофруктокиназы, замедление гликолиза и переключение гликолиза на глюконеогенез.
| Включение субстратов в глюконеогенез
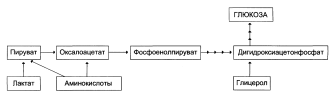 Главные пути глюконеогенсза в печени
Главные пути глюконеогенсза в печени
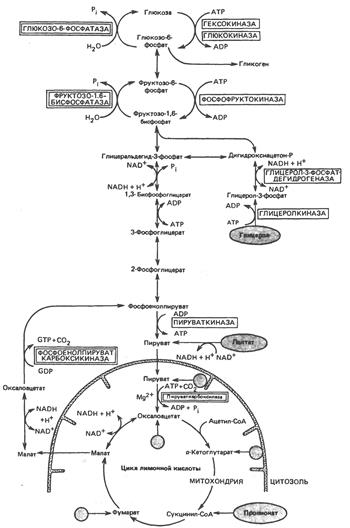 Кружок со стрелкой показывает места вступления в цикл глюкогенных аминокислот после переаминирования. Названия ключевых ферментов глюконсогенеза заключены в двойной прямоугольник. АТР, необходимый для поддержания глюконеогенеза, генерируется в ходе окисления ацетил-СоА, который в свою очередь образуется из длинноцепочечных жирных кислот или из лактата (через пируват. при действии пируватдегидрогеназы). У жвачных животных важным источником ацстил-СоА служит пропионат.
Регуляция реакций второго субстратного цикла фруктозо-2,6-бисфосфатом:
Кружок со стрелкой показывает места вступления в цикл глюкогенных аминокислот после переаминирования. Названия ключевых ферментов глюконсогенеза заключены в двойной прямоугольник. АТР, необходимый для поддержания глюконеогенеза, генерируется в ходе окисления ацетил-СоА, который в свою очередь образуется из длинноцепочечных жирных кислот или из лактата (через пируват. при действии пируватдегидрогеназы). У жвачных животных важным источником ацстил-СоА служит пропионат.
Регуляция реакций второго субстратного цикла фруктозо-2,6-бисфосфатом:
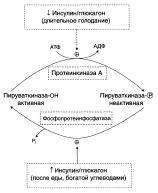
 В регуляции третьего субстратного цикла основная роль принадлежит пируваткиназе, фосфорилированная форма которой неактивна, а дефосфорилированная – активна:
В регуляции третьего субстратного цикла основная роль принадлежит пируваткиназе, фосфорилированная форма которой неактивна, а дефосфорилированная – активна:
|
| Пентозофосфатный путь
Пентозофосфатный цикл (фосфоглюконатный путь) не приводит к синтезу АТР, он выполняет две главные функции:
1) образование NADPH для восстановительных синтезов, таких, как синтез жирных кислот и стероидов;
2) обеспечение рибозой синтеза нуклеотидов и нуклеиновых кислот. Недостаточность ряда ферментов пентозофосфатного пути является причиной гемолиза эритроцитов. Например, одна из форм гемолитической анемии обусловлена недостаточностью глюкозо-6-фосфатдегидрогеназы.
Метаболическое значение пентозофосфатного пути
Сравнение с гликолизом
Пентозофосфатный путь существенно отличается от гликолиза. Окисление осуществляется на первой стадии, и в нем участвует не NAD, как в гликолизе, a NADP; одним .из продуктов пентозофосфатного пути является СО2, который в реакциях гликолиза не образуется. Наконец, пентозофосфатный путь не генерирует АТР.
Образование восстановительных эквивалентов
Значение метаболического пути для различных тканей можно оценить по его активности. Пентозофосфатный путь активно протекает в печени, жировой ткани, коре надпочечников, щитовидной железе, эритроцитах, семенниках и в молочных железах в период лактации; он неактивен в нелактирующей молочной железе и малоактивен в скелетных мышцах. Все ткани, в которых активность данного пути высока, используют в реакциях восстановительного синтеза NADPH, например в реакциях синтеза жирных кислот, стероидов, аминокислот (с участием глутаматдегидрогеназы) или восстановленного глутатиона в эритроцитах. Вероятно, в условиях активного липогенеза или при наличии любой системы, утилизирующей NADPH, возрастает активная деградация глюкозы по пентозофосфатному пути в связи с увеличением отношения NADP: NADPH. В условиях, которые возникают после приема пищи, может индуцироваться синтез глюкозо-6-фосфат-дегидрогеназы и 6-фосфоглюконатдегидрогеназы.
Образование рибозы
Пентозофосфатный путь поставляет рибозу для синтеза нуклеотидов и нуклеиновых кислот. Источником рибозы является интермедиат рибозо-5-фосфат, который в реакции с АТР образует PRPP, используемый в биосинтезе нуклеотидов. Мышечная ткань содержит очень малые количества глюкозо-6-фосфатдегидрогеназы и 6-фосфоглюконатдегидрогеназы. Тем не менее скелетная мышца способна синтезировать рибозу. Вероятно, это осуществляется при обращении неокислительной фазы пентозофосфатного пути, утилизирующей фруктозо-6-фосфат. Таким образом, синтез рибозы может осуществляться в ткани, если в ней протекает часть реакций пентозофосфатного пути.
|
Последовательность реакций пентозофосфатного пути:
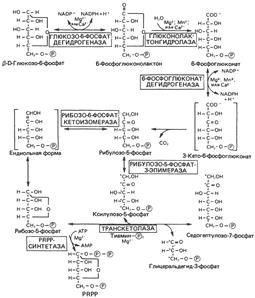
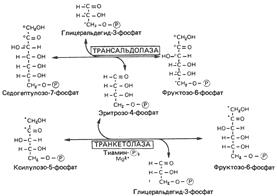 PRPP – 5-фосфорибозил-1-пирофосфат.
PRPP – 5-фосфорибозил-1-пирофосфат.
|