Лекция 8. Углеводы. Обмен углеводов.
Строение, свойства и функции основных моно-, олиго- и полисахаридов. Общие пути обмена глюкозы в клетке.
Синтез и распад гликогена. Механизм ветвления гликогена. Ковалентная модификация и аллостерическая регуляция гликогенфосфорилазы и гликогенсинтазы. Механизм синхронизации мышечного сокращения и гликогенолиза. Гликогенозы.
Гликолиз: последовательность реакций. Гликолитическая оксидоредукция. Субстратное фосфорилирование.
Ключевые реакции глюконеогенеза. Аллостерическая регуляция ферментов гликолиза и глюконеогенеза. Роль фруктозо-2,6-бисфосфата.
Реакции пентозофосфатного пути превращения глюкозы. Образование восстановительных эквивалентов и рибозы.
Метаболизм фруктозы и галактозы. Регуляция уровня глюкозы в крови. Источники глюкозы крови. Цикл Кори и глюкозо-аланиновый цикл. Почечный порог для глюкозы, глюкозурия. Толерантность к глюкозе.
| Краткое содержание | Конспект |
Синтез и распад гликогена. Механизм ветвления гликогена. Ковалентная модификация и аллостерическая регуляция гликогенфосфорилазы и гликогенсинтазы. Механизм синхронизации мышечного сокращения и гликогенолиза. Гликогенозы
 Образование УДФ-глюкозы
Образование УДФ-глюкозы
 Образованная УДФ-глюкоза далее используется как донор остатка глюкозы при синтезе гликогена. Эту реакцию катализирует фермент гликогенсинтаза (глюкозилтрансфераза).
Поскольку гликоген в клетке никогда не расщепляется полностью, синтез гликогена осуществляется путём удлинения уже имеющейся молекулы полисахарида, называемой «затравкой», или «праймер». К «затравке» последовательно присоединяются молекулы глюкозы. В состав «затравки» может входить белок гликогенин, в котором к ОН-группе одного из тирозиновых остатков присоединена олигосахаридная цепочка, состоящая из 8 остатков глюкозы.
Разветвлённая структура гликогена образуется при участи фермента амило-1,4→1,6-гликозилтрансферазы (фермент ветвления).
Распад гликогена (гликогенолиз)
Стадией, лимитирующей скорость гликогенолиза, является реакция, катализируемая фосфорилазой:
(С6)n + Pi → (С6)n-1 + Глюкозо-1-фосфат
Гликоген Гликоген
Фермент специфично катализирует фосфоролитическое расщепление (фосфоролиз) (1→4)-связей гликогена, продуктом является глюкозо-1-фосфат. Остатки глюкозы отщепляются от дальних концов молекулы гликогена до тех пор, пока на ветвях, идущих от точки ветвления ([1→6]-связи), не останется примерно по 4 остатка глюкозы. Другой фермент (α-[1→4] → α-[1→4]-глюкантрансфераза) переносит трисахаридный фрагмент с одной цепи на другую, экспонируя (1→6)-пункт ветвления. Гидролитическое расщепление (1→6)-связей осуществляет деветвящий (дебранчинг) фермент (амило-[1→6]-глюкозидаза); по-видимому, это второй вид активности глюкантрансферазы. После удаления ветви на цепь гликогена снова действует фосфорилаза. Совместное действие фосфорилазы и других рассмотренных выше ферментов приводит к полному распаду гликогена. Реакция, катализируемая фосфоглюкомутазой, обратима; поэтому из глюкозо-1-фосфата может образовываться глюкозо-6-фосфат.
В печени и почках (но не в мышцах) имеется специфический фермент глюкозо-6-фосфатаза, отщепляющий фосфат от глюкозо-6-фосфата. Образовавшаяся глюкоза диффундирует из клеток в кровь. Это заключительная стадия гликогенолиза в печени, приводящего к повышению содержания глюкозы в крови.
Ковалентная модификация и аллостерическая регуляция гликогенфосфорилазы и гликогенсинтазы
Образованная УДФ-глюкоза далее используется как донор остатка глюкозы при синтезе гликогена. Эту реакцию катализирует фермент гликогенсинтаза (глюкозилтрансфераза).
Поскольку гликоген в клетке никогда не расщепляется полностью, синтез гликогена осуществляется путём удлинения уже имеющейся молекулы полисахарида, называемой «затравкой», или «праймер». К «затравке» последовательно присоединяются молекулы глюкозы. В состав «затравки» может входить белок гликогенин, в котором к ОН-группе одного из тирозиновых остатков присоединена олигосахаридная цепочка, состоящая из 8 остатков глюкозы.
Разветвлённая структура гликогена образуется при участи фермента амило-1,4→1,6-гликозилтрансферазы (фермент ветвления).
Распад гликогена (гликогенолиз)
Стадией, лимитирующей скорость гликогенолиза, является реакция, катализируемая фосфорилазой:
(С6)n + Pi → (С6)n-1 + Глюкозо-1-фосфат
Гликоген Гликоген
Фермент специфично катализирует фосфоролитическое расщепление (фосфоролиз) (1→4)-связей гликогена, продуктом является глюкозо-1-фосфат. Остатки глюкозы отщепляются от дальних концов молекулы гликогена до тех пор, пока на ветвях, идущих от точки ветвления ([1→6]-связи), не останется примерно по 4 остатка глюкозы. Другой фермент (α-[1→4] → α-[1→4]-глюкантрансфераза) переносит трисахаридный фрагмент с одной цепи на другую, экспонируя (1→6)-пункт ветвления. Гидролитическое расщепление (1→6)-связей осуществляет деветвящий (дебранчинг) фермент (амило-[1→6]-глюкозидаза); по-видимому, это второй вид активности глюкантрансферазы. После удаления ветви на цепь гликогена снова действует фосфорилаза. Совместное действие фосфорилазы и других рассмотренных выше ферментов приводит к полному распаду гликогена. Реакция, катализируемая фосфоглюкомутазой, обратима; поэтому из глюкозо-1-фосфата может образовываться глюкозо-6-фосфат.
В печени и почках (но не в мышцах) имеется специфический фермент глюкозо-6-фосфатаза, отщепляющий фосфат от глюкозо-6-фосфата. Образовавшаяся глюкоза диффундирует из клеток в кровь. Это заключительная стадия гликогенолиза в печени, приводящего к повышению содержания глюкозы в крови.
Ковалентная модификация и аллостерическая регуляция гликогенфосфорилазы и гликогенсинтазы
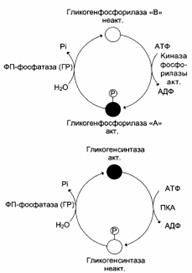
| Кружками обозначены молекулы фермента: активные – чёрные, неактивные – белые. ФП – фосфатаза (ГР) – фосфопротеин-фосфатаза гранул гликогена |
Синтез и распад гликогена
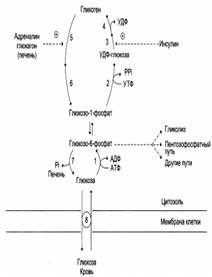
| 1 – гексокиназа или глюкокиназа (печень); 2 – УДФ-глюклпирофосфорилаза; 3 – гликогенсинтаза; 4 – амило-1,4→1,6-гликозилтрансфераза (фермент ветвления); 5 – гликогенфосфорилаза; 6 – «деветвящий» фермент; 7 – глюкозо-6-фосфатаза (печень); 8 – транспортные системы ГЛЮТ. |
Гликогенозы (болезни, связанные с накоплением гликогена)
Термин «гликогеноз» является общим для группы наследственных заболеваний, характеризующихся отложением в тканях либо ненормально больших количеств гликогена, либо необычных его видов.
При гликогенозе I типа (болезнь Гирке) клетки печени и извитых почечных канальцев заполнены гликогеном, однако эти запасы оказываются недоступными: об этом свидетельствует гипогликемия, а также отсутствие повышения уровня глюкозы в крови в ответ на адреналин и глюкагон. Обычно у этих больных развиваются кетоз и гиперлипемия, что вообще характерно для состояния организма при недостатке углеводов. В печени, почках и тканях кишечника активность глюкозо-6-фосфатазы либо крайне низка, либо вообще отсутствует.
Гликогеноз II типа (болезнь Помпа) ведет к фатальным последствиям и характеризуется отсутствием лизосомальной α-(1→4)- и (1→6)-глюкозидазы (кислой мальтазы), функцией которой является деградация гликогена, предотвращающая его накопление в лизосомах.
Гликогеноз III типа (лимитдекстриноз; болезнь Форбса или болезнь Кори) характеризуется отсутствием деветвящего фермента; в результате накапливается характерный разветвленный полисахарид (остаточный декстрин).
Гликогеноз IV типа (амилопектиноз; болезнь Андерсен) характеризуется отсутствием ветвящего фермента, в результате чего накапливается полисахарид, содержащий незначительное число ветвей. Обычно летальный исход наступает из-за сердечной или печеночной недостаточности в первый год жизни.
Отсутствие мышечной фосфорилазы (миофосфорилазы) является причиной гликогеноза V типа (миофсофорилазная недостаточность; синдром Мак-Ардля). У больных наблюдается пониженная выносливость к физическим нагрузкам. Хотя в их скелетных мышцах имеется аномально высокое содержание гликогена2,5-4,1%), в крови после выполнения физической работы почти или вообще не обнаруживается лактат.
Описаны гликогенозы, связанные с недостаточностью фосфорилазы в печени (гликогеноз VI типа), недостаточностью фосфофруктокиназы в мышцах и эритроцитах (гликогенез VII типа; болезнь Таруи), а также гликогеноз, обусловленный недостаточностью киназы фосфорилазы. Сообщалось также о случаях недостаточности аденилаткиназы и сАМР-зависимой протеинкиназы.
Катаболизм гликогена
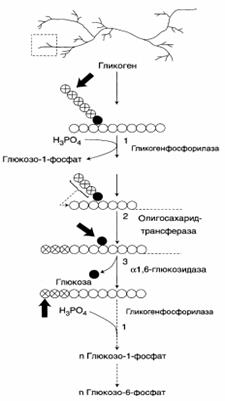
| В рамке – фрагмент гликогена с точкой ветвления. Закрашенный кружок – глюкозный остаток, связанный α-1,6-гликозидной связью. Светлые и заштрихованные кружки – глюкозные остатки в линейных участках и боковых ветвях, связанные α-1,4-гликозидной связью. 1 – гликогенфосфорилаза; 2 – олигосахарид-трансфераза; 3 – α-1,6-глюкозидаза. |
Мобилизация гликогена
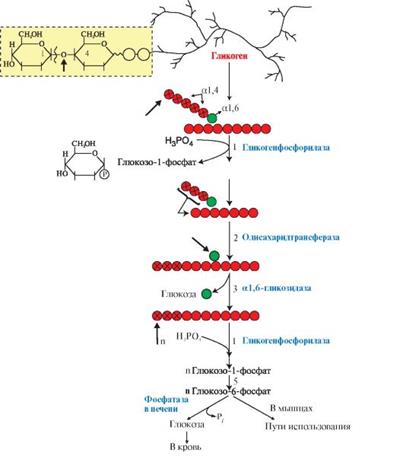
Зелёные кружочки – глюкозный остаток, связанный α1,6-гликозидной связью;
Красные кружки – гликозидные остатки в линейных участках и боковых ветвях, связанные α1,4-гликозидной связью.
1 – реакции фосфоролиза – отщепление глюкозного остатка в форме глюкозо-1-фосфата. В реакции используется неорганический фосфат;
2 – перенос трёх мономеров, оставшихся неотщеплёнными до точки ветвления, на нередуцирующий конец цепи и удлинение этой цепи;
3 – отщепление глюкозного остатка в точе ветвления (гидролиз α1,6-гликозидной связи) и образование свободной глюкозы;
4 – повторяющееся действие гликогенфосфорилазы и других ферментов с образованием основного продукта – глюкозо-1-фосфата;
5 – образование глюкозо-6-фосфата и использование его в печени и мышцах.
Инозитолфосфатный механизм регуляции синтеза и распада гликогена в печени адреналином и Са2+
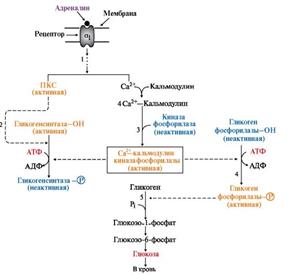
1 - взаимодействие адреналина с α1-рецептором передает сигнал через инозитолфосфатную систему. Это сопровождается активацией фосфолипазы С, мобилизацией Сa2+ из ЭР и активацией протеинкиназы С (ПКС).
2 - протеинкиназа С фосфорилирует гликогенсинтазу и переводит ее в неактивное состояние.
3 - комплекс 4Са2+-кальмодулин активирует киназу фосфорилазы и кальмодулинзависимые протеинкиназы.
4 - киназа фосфорилазы фосфорилирует гликогенфосфорилазу и тем самым ее активирует.
5 - гликогенфосфорилаза катализирует первую реакцию распада гликогена.
Реакции пентозофосфатного пути превращения глюкозы. Образование восстановительных эквивалентов и рибозы.
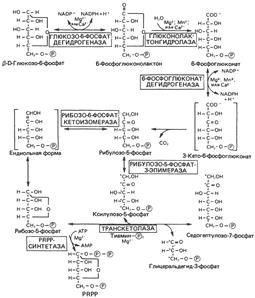
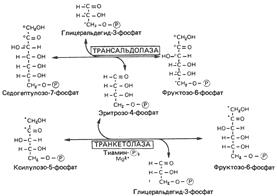 PRPP – 5-фосфорибозил-1-пирофосфат.
PRPP – 5-фосфорибозил-1-пирофосфат.
Лекция 8. Углеводы. Обмен углеводов.
Дата: 2018-11-18, просмотров: 1135.