Текучесть мембран, влияние на нее жирнокислотного состава мембранных липидов, поливалентных катионов, холестерола. Мембранные белки: интегральные и периферические. Асимметрия мембран. Сборка мембран
Поможем в ✍️ написании учебной работы
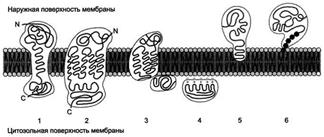 Сборка мембран
Полярные липиды, к числу которых относятся фосфоглицеролы, сфинголипиды и гликолипиды, не запасаются в жировых клетках, а встраиваются в клеточные мембраны, причем в строго определенных соотношениях. Фосфоглицеролы, синтезируемые ферментами плазматического ретикулума, встраиваются в основном в липидный бислой ретикулума. Общая площадь эндоплазматического ретикулума особенно велика в клетках печени и поджелудочной железы. Мембраны эндоплазматического ретикулума служат предшественниками мембран аппарата Гольджи. От аппарата Гольджи постоянно отшнуровываются мембранные пузырьки, в которых продукты секреции транспортируются к плазматической мембране. Эти пузырьки часто сливаются с плазматической мембраной. Фосфоглицеролы могут переноситься из эндоплазматического ретикулума в митохондрии также при помощи транспортных белков. Таким образом, в клетке существует поток вновь синтезированных полярных липидов, направленный к различным типам клеточных мембран.
Сборка мембран
Полярные липиды, к числу которых относятся фосфоглицеролы, сфинголипиды и гликолипиды, не запасаются в жировых клетках, а встраиваются в клеточные мембраны, причем в строго определенных соотношениях. Фосфоглицеролы, синтезируемые ферментами плазматического ретикулума, встраиваются в основном в липидный бислой ретикулума. Общая площадь эндоплазматического ретикулума особенно велика в клетках печени и поджелудочной железы. Мембраны эндоплазматического ретикулума служат предшественниками мембран аппарата Гольджи. От аппарата Гольджи постоянно отшнуровываются мембранные пузырьки, в которых продукты секреции транспортируются к плазматической мембране. Эти пузырьки часто сливаются с плазматической мембраной. Фосфоглицеролы могут переноситься из эндоплазматического ретикулума в митохондрии также при помощи транспортных белков. Таким образом, в клетке существует поток вновь синтезированных полярных липидов, направленный к различным типам клеточных мембран.
| Молекулы различных полярных липидов после завершения их синтеза встраиваются в липидный бислой клеточных мембран в определенных соотношениях. Основная масса полярных липидов встраивается в бислой мембран эндоплазматического ретикулума. Эти липиды поступают затем последовательно в мембраны аппарата Гольджи, секреторные пузырьки и плазматическую мембрану. При помощи специфических белков-переносчиков липиды эндоплазматического ретикулума переносятся через цитозоль и встраиваются в митохондриальные и ядерные мембраны. | 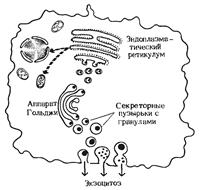
|
Микротранспорт: пассивный транспорт (простая и облегченная диффузия), активный транспорт (первичный и вторичный). Унипорт и котранспорт (симпорт и антипорт). Макротранспорт: эндоцитоз (пиноцитоз и фагоцитоз) и экзоцитоз. Окаймленные ямки и пузырьки. Роль клатрина
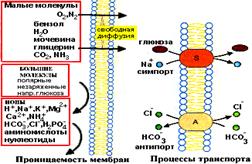 Перенос вещества и информации через мембраны:
1. Трансмембранное перемещение малых молекул:
- Диффузия (пассивная и облегченная).
- Активный транспорт.
2. Трансмембранное перемещение крупных молекул:
- Эндоцитоз.
- Экзоцитоз.
3. Передача сигнала через мембраны
Рецепторы клеточной поверхности
1. Передача сигнала (например, глюкагон→цАМФ).
2. Интернализация сигнала (сопряженная с эндоцитозом, например рецептор ЛПНП).
4. Межклеточные контакты и коммуникации
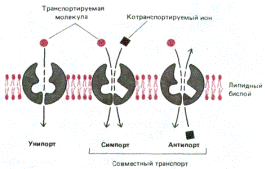
Схема работы белков-переносчиков, функционирующих по принципу унипорта, симпорта и антипорта:
Перенос вещества и информации через мембраны:
1. Трансмембранное перемещение малых молекул:
- Диффузия (пассивная и облегченная).
- Активный транспорт.
2. Трансмембранное перемещение крупных молекул:
- Эндоцитоз.
- Экзоцитоз.
3. Передача сигнала через мембраны
Рецепторы клеточной поверхности
1. Передача сигнала (например, глюкагон→цАМФ).
2. Интернализация сигнала (сопряженная с эндоцитозом, например рецептор ЛПНП).
4. Межклеточные контакты и коммуникации
Схема работы белков-переносчиков, функционирующих по принципу унипорта, симпорта и антипорта:
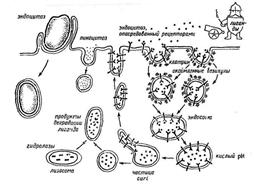
 Виды макротранспорта (эндоцитоз, экзоцитоз):
Виды макротранспорта (эндоцитоз, экзоцитоз):
 Строение и функционирование Nа+,К+-АТФ-азы плазматической мембраны:
Строение и функционирование Nа+,К+-АТФ-азы плазматической мембраны:
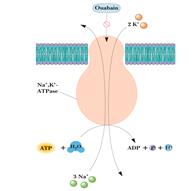 По какому принципу работает Nа+,К+-АТФ-аза?
По какому принципу работает Nа+,К+-АТФ-аза?
Дата: 2018-11-18, просмотров: 1828.
Материалы представленные на сайте исключительно с целью ознакомления пользователям Интернета и не преследуют коммерческих целей или нарушение авторских прав.
© 2018 - 2026