Механизм действия NO
Низкомолекулярный газ NO легко проникает через клеточные мембраны и компоненты межклеточного вещества, однако время его полужизни (в среднем не более 5с) и расстояние возможной диффузии (небольшое, в среднем 30 мкм) ограничиваются высокой реакционной способностью молекулы и ее взаимодействием со многими возможными субстратами [2].
Действие, оказываемое NO на клетки, во многом зависит от количества газа. В небольших количествах, продуцирующихся обычно конститутивными формами NO-синтазы, эффект NO в основном связан с влиянием на гемовую группу растворимой (цитозольной) формы гуанилатциклазы. Активированный фермент синтезирует циклический гуанозин монофосфат (цГМФ) – активный внутриклеточный посредник, регулирующий работу мембранных ионных каналов, процессы фосфорилирования белков (через протеинкиназы), активность фосфодиэстеразы, а также др. реакции [2].
В больших концентрациях, образующихся, как правило, индуцибельной изоформой NO-синтазы, NO может оказывать на клетки токсический эффект, связанный как с прямым действием на железосодержащие ферменты, так и с образованием сильного окислителя, очень реакционного и токсичного свободнорадикального соединения пероксинитрита [14]. Пероксинитрит (ONOO-) образуется при взаимодействии NO с радикальным супероксид анионом(О2-):
NO + O2- = ONOO- [2]
Участие NO в защитных иммунологических реакциях
Участие нитропроизводных (производных нитритов и нитратов) во многих патологических процессах, в том числе и опухолевом росте, было известно давно. Первоначально разрозненные и часто необъяснимые данные о связи противомикробного и противоопухолевого действия макрофагов и нитропроизводных прояснились после открытия синтеза NO в эндотелиальных клетках. Действие макрофагов на чужеродные агента также стали связывать с NO, и многочисленные эксперименты подтвердили, что макрофаги способны синтезировать NO-синтазу и выделять большое количество газа [2]. Уже отмечалось, что NO-синтаза макрофагов является индуцибельным ферментом. В нормальных условиях клетки не содержат этот фермент и не продуцируют NO [10]. Под влиянием липополисахаридов микробного происхождения или цитокинов – высокоактивных межклеточных посредников, выделяющихся, в частности, лимфоцитами при их контакте с чужеродными агентами, в макрофагах начинается синтез индуцибельной изоформы NO-синтазы, образующей большой объем NO, оказывающего, в свою очередь, цитостатическое и цитолитическое действие на бактериальные и чужеродные (в том числе и раковые) клетки [2].
Нейтрофилы также способны экспрессировать индуцибельную форму NO-синтазы и синтезировать NO, однако данные о цитотоксическом действии этих клеток, связанном с NO, неизвестны.
Известно, что нейтрофилы и макрофаги способны активно образовывать свободные радикалы кислорода, и, возможно, образование пероксинитрита в реакции NO со свободными радикалами может усиливать антимикробный эффект этих клеток [2].
NO и кровеносные сосуды
Значение NO в кровоснабжении многогранно. Прежде всего NO – мощный сосудорасширяющий агент. Эндотелий постоянно продуцирует небольшие количества NO (так называемый базовый фон), а при различных воздействиях – механических (например, при усилении тока или пульсации крови), химических бактериальных и вирусных – синтез NO в эндотелиальных клетках значительно повышается [2].
Расширение сосудов связано с диффузией NO из эндотелия к соседним гладкомышечным клеткам стенки сосуда, активацией в них гуанилатциклазы и образованием цГМФ [8]. Повышение уровня цГМФ приводит к снижению уровня ионов кальция в цитозоле клеток и ослаблению связи между миозином и актином, что и позволяет клеткам расслабиться, то есть принять первоначальную форму и размеры. Следует помнить, что расслабление мышечных клеток обусловлено не внутриклеточными процессами, а связано с внешними по отношению к клеткам механическим факторам, в частности для гладких мышц сосудов это упругость эластических волокон, окружающих и оплетающих гладкомышечные клетки и растягивающих клетки после прекращения процесса сокращения и устранения связи между актином и миозином [2].
Действует NO очень быстро – образование цГМФ происходит через 5с, а начало расслабления гладких мышц – через 10с после добавления NO в культуру изолированных кровеносных сосудов. Открытие сосудорасширяющего действия NO позволило прояснить механизм действия самого распространенного и эффективного лекарственного средства, применяемого для лечения спазма коронарных артерий – нитроглицерина. При расщеплении препарата образуется NO, приводящий к расширению сосудов сердца и снимающий в результате этого чувство боли [2].
Большое значение NO имеет в регуляции мозгового кровообращения. Известные более ста лет назад данные об усилении кровотока в активно работающих областях мозга получили после открытия сосудорасширяющего действия NO более полную интерпретацию. Имеется несколько источников NO для регуляции просвета мозговых сосудов [8]. Это эндотелий сосудов, нейроны, содержащие NO-синтазу и оплетающие своими отростками стенки сосудов и астроциты, образующие периваскулярные оболочки (рис. 3). Активация нейронов какой-либо области мозга приводит к возбуждению нейронов, содержащих NO-синтазу, и/или астроцитов, в которых также может индуцироваться синтез NO, и выделяющийся из клеток газ приводит к локальному расширению сосудов в области возбуждения [2].
С NO связывает и развитие септического шока, когда большое количество микробов, циркулирующих в крови, резко активирует синтез газа в эндотелии, что приводит к длительному и сильному расширению мелких кровеносных сосудов и как следствие – значительному снижению артериального давления, с трудом поддающемуся терапевтическому воздействию [2].
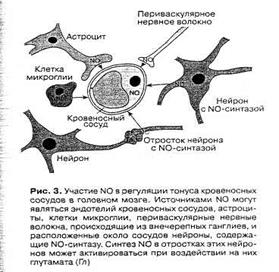
Рис. 5. Участие NO в регуляции тонуса кровеносных сосудов в головном мозге [2].
NO, образующийся в эндотелии, оказывает влияние и на взаимодействие клеток крови с эндотелием. Газ препятствует прилипанию лейкоцитов и кровяных пластинок к эндотелию и также снижает агрегацию последних [9]. Такое действие NO может иметь большое значение на ранних стадиях развития тромбов и в генезе атеросклеротических повреждений стенки сосудов. Участие NO в развитии атеросклероза может заключаться и еще в одной стороне его действия. NO может выступать в роли антиростового фактора, препятствующего пролиферации гладкомышечных клеток стенки сосудов, важного звена в патогенезе болезни [2].
NO и нервная система
В нервной системе NO имеет большое значение, как в нормальных физиологических условиях, так и при различной патологии. Источниками NO в ЦНС являются нейроны, нейроглиальные клетки – астроциты и клетки микроглии и эндотелий кровеносных сосудов [2].
Нейроны, содержащие NO-синтаза, находятся во многих отделах ЦНС и большинстве изученных периферических ганглиев нервной системы. В коре больших полушарий в среднем 2% нейронов содержат NO-синтазу, в большинстве отделов головного мозга число таких нейронов также невелико. Однако имеются области и с высоким их содержанием. Так, максимальное количество нейронов, содержащих NO-синтазу, находится в коре мозжечка, где большинство клеток-зерен и корзинчатых нейронов содержат фермент [8]. Самые крупные нейроны коры мозжечка – клетки Пуркинье – не содержат NO-синтазу. Сравнительно много нейронов, содержащих NO-синтазу, находится в обонятельных луковицах, а также в некоторых отделах гиппокампа и полосатого тела. Только в мозжечке нейроны, содержащие NO-синтазу, составляют компактную популяцию клеток, а в остальных отделах – это одиночные, редко расположенные клетки (рис. 4) [2].

Рис.6. Нейроны, содержащие NO -синтазу, в коре больших полушарий белой крысы [2].
Обобщая имеющиеся данные о нейронах ЦНС, содержащих NO-синтазу, следует отметить, что преимущественная часть их относится к небольшим по размерам клеткам, многие из которых не содержат дендритных шипиков и являются ассоциативными нейронами. Крупные клетки, например пирамидные нейроны коры больших полушарий или гиппокампа, моторные нейроны передних рогов спинного мозга, не содержат NO-синтазу. NO-синтаза сосуществует в нервных клетках с другими традиционными нейромедиаторами и нейропептидами, чаще фермент определяется в холинэргических нейронах [2].
Большое внимание уделяется NO в реализации нервных воздействий на ткани внутренних органов. Нервы, содержащие NO-синтазу, показаны практически во всех изученных внутренних органах, преимущественно в стенке кровеносных сосудов, где они, наряду с эндотелием, могут оказывать сосудорасширяющий эффект [8] NO признается одним из основных эффекторных агентов в так называемых неадренэргических-нехолинэргических нервах. В периферических нервах подробно изучено сосуществование NO-синтазы с нейропептидами. Наиболее часто фермент определяется вместе с вазоактивным кишечнымнейропептидом (VIP) и нейропептидом Y [2].
Значение NO в ЦНС в нормальных условиях связывают с тремя процессами (так называемая NO-гипотеза):
1) Участие в межнейронной связи в качестве своеобразного нейромедиатора, причем основное значение, как полагают, NO имеет в синаптической пластичности, под которой понимают эффективность синаптической передачи;
2) Регуляция церебрального кровотока;
3) Установление межнейронных синаптических взаимосвязей во время развития нервной системы[2].
NO как нейромедиатор
Нейромедиаторная сущность NO заключается в том, что оно синтезируется при возбуждении нейрона (в ответ на поступление ионов кальция) и, диффундируя в соседние клетки, активизирует в них образование цГМФ, способного влиять на проводимость ионных каналов и, таким образом, изменять электрогенез нейронов [2]. NO отличается от традиционных нейромедиаторов тем, что он оказывает воздействие на ионные каналы не через плазмалеммальные рецепторы, а изнутри, со стороны цитоплазмы. Кроме того, действие NO не ограничивается только областью синаптических контактов, газ может влиять на ионные каналы на значительной площади плазматической мембраны нейрона [11].
Участие NO в синаптической пластичности наиболее ярко проявляется в таких процессах, как длительная синаптическая потенция (повышение эффективности проведения возбуждения через синапс для каждого последующего импульса в их последовательности), более детально проанализированная в гиппокампе, и длительная синаптическая депрессия (снижение эффективности проведения возбуждения через синапс), лучше исследованная в коре мозжечка[2].
В первом случае возбуждение постсинаптического окончания приводит к повышению внутриклеточного уровня Ca2+/кальмодулина, активирующих NO-синтазу и образование NO. NO, диффундируя в пресинаптическое окончание, вызывает образование цГМФ, что приводит к усилению и увеличению длительности выделения из пресинапса нейромедиатора, который оказывает возбуждающее действие на постсинаптический нейрон, и таким образом возникает обратная положительная связь с постоянным усилением. Такому механизму особое влияние уделяется в синапсах с наиболее распространенными и сильновозбуждающими нейромедиаторами ЦНС глутаматом и особому глутаматному рецептору – NMDA- рецептору, являющемуся трансмембранным каналом для ионов кальция (рис 7) [2].
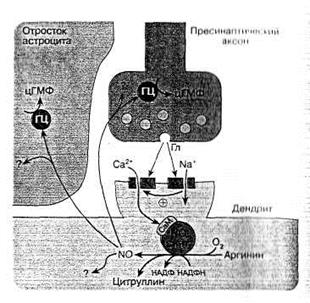
Рис. 7. Схема возбуждающего глутаматного синапса ЦНС и возможное участие NO в его деятельности [2].
Выделяющийся из синаптических пузырьков возбуждающий нейромедиатор глутамат (Гл) влияет на АМРА (А)-рецепторы, что приводит к деполяризации плазматической мембраны и открытию NMDA(N)-каналов (последнее связано с высвобождением вследствие изменения потенциала мембраны из ионного канала ионов магния, которые закрывали канал). Через NMDA-каналы в клетку поступает большой объем ионов кальция, активирующих конститутивную изоформу NO-синтазы. Образующийся NO, диффундируя в пресинаптический аксон, активирует в нем гуанилатциклазу (ГЦ) и синтез цГМФ, последний способствует усилению выделению нейромедиатора глутамата, что повышает эффективность синаптической передачи (феномен длительной синаптической потенции). NO может также влиять на окружающие астроциты и активировать в них гуанилатциклазу.
Дополнительные обозначения: СаМ – кальмодулин; ? – возможное влияние NO на другие не связанные с гуанилатциклазой клеточные системы нейронов и астроцитов.
С длительной синаптической потенцией, прежде всего в гиппокампе, связывают пластичность межнейронных связей, лежащих в основе памяти. Такое предположение основывается на ставших уже классическими представлениях Д.Хэбба (1949) о повышении эффективности синаптической передачи при возбужденном состоянии постсинаптического нейрона. Физиологические наблюдения об участии NO в процессах памяти и обучения противоречивы. Наряду с работами, в которых показано нарушение процессов обучения у экспериментальных животных при введении ингибиторов NO-синтазы, имеются и исследования с противоположными результатами. Неоднозначность получаемых результатов, возможно, связана с тем, что вводимые в организм ингибиторы NO-синтазы оказывают действие не только на всю нервную систему, но и на все органы и ткани животных, что не позволяло достичь локального влияния на продукцию NO в мозге [2].
В переживающих (живущих некоторое время в условиях культуры тканей) срезах головного мозга был показан и другой механизм действия NO на пресинаптические окончания. NO способен инициировать выделение нейромедиатора дофамина из нейронов не посредством экзоцитоза, а путем трансмембранной диффузии при участии особого мембранного переносчика дофамина, участвующего в его захвате из межклеточной среды [8]. В этом процессе не участвует цГМФ и предполагается прямое влияние NO на транспортные белки мембран [2].
Длительную синаптическую депрессию, в частности в контактах между параллельными волокнами, являющимися аксонами самых маленьких нейронов в организме человека, так называемых клеток-зерен, и нейронами Пуркинье в коре мозжечка, связывают с десенсибилизацией другого класса глутаматных рецепторов, так называемых АМРА-рецепторов. Выделяющийся из аксонов корзинчатых нейронов NO диффундирует к нейронам Пуркинье и активирует в них синтез цГМФ, что приводит к инактивации АМРА-рецепторов и снижению эффективности работы синапсов между клетками-зернами и нейронами Пуркинье [13].
В основе первого положения NO-гипотезы (см. выше) лежит возможность образования NO в постсинаптическом окончании при его возбуждении влияние газа на пресинаптический аксон. Однако за счет диффузии к соседним нервным клеткам NO может оказывать влияние не только на пресинаптическое расширение аксона, формирующего синапс на этом постсинаптическом окончании, но и на близлежащие аксоны и дендриты, модулируя их активность (рис. 6) [2].
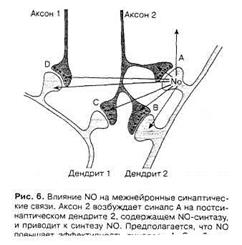
Рис.8. Влияние NO на межнейронные синаптические связи [2].
Традиционная формулировка положения об участии NO в межнейронной коммуникации ограничивается обычно возможностью синтеза и выделения NO из локальной области нейрона – постсинаптического окончания. Однако, как показывают результаты свето- и электронно-микроскопических исследований, NO-синтаза определяется во всем объеме тела нейронов – в перикарионе, аксоне и дендритах. Поскольку при возбуждении нейрона по всей длине его отростков и в теле уровень кальция циклически колеблется (образуются своеобразные кальциевые волны), можно считать, что синтез и выделение NO могут инициироваться в любом участке тела и отростков нейронов. Таким образом, нейроны, содержащие NO-синтазу, способны создавать вокруг себя поле воздействия, то есть могут считаться своеобразными полевыми нейронами в отличие от традиционных нейронов, связанных друг с другом в локальных участках – синапсах [2].
Основное внимание в процессах синаптической пластичности уделяется нейронам, однако нельзя не учитывать и роль глии. Известно, что астроциты способны продуцировать NO, причем они обладают как конститутивной (в небольшом количестве), так и индуцибельной NO-синтазой [8]. Если принять во внимание, что число астроцитов в 10-100 раз превосходит (в зависимости от области мозга) количество нейронов, то их роль в продукции NO и влиянии на механизмы электрогенеза нейронов может являться весьма значимой [2].
Значение астроцитов как источника NO особенно ярко проявляется при патологии ЦНС. При многих нейродегенеративных заболеваниях, ишемии, травмах, опухолях головного мозга астроциты начинают экспрессировать NO-синтазу и продуцировать большой объем NO. С этим связывают гибель нейронов и других макроглиальных клеток, в частности олигодендроцитов [2].
Дата: 2019-12-10, просмотров: 387.