БИОЛОГИЯ И МИКРОБИОЛОГИЯ
РАЗДЕЛ 1 БИОЛОГИЯ
Лекция № 1 Современный этап развития биологии
Лекция № 2 Общая характеристика живого
Свойства живых организмов
2.1 Обмен веществ. Все живые организмы обладают способностью извлекать, преобразовывать и использовать энергию окружающей среды либо в виде питательных веществ, либо в форме солнечного излучения. Во внешнюю среду они возвращают продукты распада и преобразованную энергию в виде тепла. То есть организмы способны к обмену веществом и энергией с окружающей средой.
Обмен веществ является одним из существенных критериев жизни. Это свойство отражено в определении жизни, которое сформулировал Ф.Энгельс более ста лет назад:
«Жизнь есть способ существования белковых тел, существенным моментом которого является постоянный обмен веществ с окружающей их внешней средой, причем с прекращением этого обмена веществ прекращается и жизнь, что приводит к разложению белка».
В это определение вошли два важных положения:
А) жизнь тесно связана с белковыми веществами;
Б) непременным условием жизни является постоянный обмен веществ, с прекращением которого прекращается и жизнь.
Обмен веществ белкового тела имеет две стороны:
· Пластический обмен (анаболизм) – это совокупность реакций, обеспечивающих построение клетки и обновление ее состава.
· Энергетический обмен (катаболизм) – это совокупность реакций, обеспечивающих клетку энергией.
Анаболизм + катаболизм = обмен веществ (метаболизм)
Вещества, поступающие из окружающей среды в результате пластического обмена, превращаются в вещества данного организма, и из них строится тело организма. Таким образом, пластический обмен состоит из двух одновременно идущих процессов: непрерывного распада веществ – диссимиляции и непрерывного синтеза новых соединений, т.е. ассимиляции. Процессы диссимиляции и ассимиляции едины и не существуют отдельно друг от друга. В результате этих процессов живой организм все время меняется, но при этом сохраняет свою определенную структуру.
Для ассимиляции, т.е. образования нового сложного вещества, кроме «строительного материала» - разнообразных химических соединений, необходима также энергия. Эту энергию дают, в первую очередь, процессы распада, т.е. процессы диссимиляции. При этом происходит расщепление сложных органических соединений на более простые, которые окисляются до конечных продуктов, как правило, до углекислого газа и воды с выделением энергии. Все это происходит в процессе энергетического обмена – катаболизма.
Живому организму энергия требуется не только для создания новых веществ тела, но и для различных видов деятельности: работа мышц, желез, нервных клеток и др., высшим животным – для поддержания постоянной температуры тела.
Чем больше нагрузка на организм, и чем больше затрачивается энергии, тем большее количество питательных веществ должно поступать. Людям тяжелого физического труда, спортсменам при больших нагрузках необходимо усиленное питание. Несоответствие между поступающей энергией в виде питательных веществ и затрачиваемой организмом ведет к увеличению веса и заболеваниям.
Обмен веществ обеспечивает устойчивость и постоянство химического состава клетки и всего организма, а, следовательно, и их деятельность.
Динамические системы, в которых непрерывно протекают химические реакции за счет поступающих извне веществ и энергии, а продукты распада отводятся, называются открытыми системами.
Живой организм – это открытая система, т.к. он существует до тех пор, пока в него поступает пища, а также энергия из внешней среды, а некоторые продукты обмена выделяются.
Живые организмы обладают встроенной системой саморегуляции, которая поддерживает процессы жизнедеятельности и препятствует неупорядоченному распаду структур и выделению энергии. Это тесно связано с процессом обмена веществ.
Способность биологических систем противостоять изменениям и сохранять динамическое постоянство состава и свойств называется гомеостазом
Гомеостаз – относительное динамическое постоянство состава и свойств внутренней среды и устойчивость основных физиологических функций организма.
Различают: а) физиологический гомеостаз – это генетически детерминированная способность организма сохранять свой статус в изменяющихся условиях внешней среды (у млекопитающих – способность сохранять постоянство осмотического давления в клетках и рН крови);
б) гомеостаз развития - это генетически детерминированная способность организма так изменять отдельные реакции, что функции организма при этом в целом сохраняются. (У человека при удалении одной почки оставшаяся выполняет двойную нагрузку)
2.2 Способность к самовоспроизведению – это второе обязательное свойство живого.
Время жизни всех живых систем, от молекулярных структур (вирусы, прионы) до высокоорганизованных многоклеточных организмов, ограничено.
Самовоспроизведение осуществляется на всех уровнях организации живой материи – от макромолекул до организма. Благодаря этому свойству клеточные структуры, клетки и организмы сходны по строению со своими предшественниками.
В основе самовоспроизведения лежит образование новых молекул и структур, на основе информации, заложенной в нуклеиновой кислоте ДНК. Самовоспроизведение тесно связано с явлением наследственности: любое живое существо рождает себе подобных.
 Материальной основой генетических программ являются нуклеиновые кислоты: ДНК РНК белок
Материальной основой генетических программ являются нуклеиновые кислоты: ДНК РНК белок
Белок является функциональным исполнительным механизмом, который регулируется нуклеиновой кислотой. Этому соответствует одно из современных определений жизни, данное в 1965 г. советским ученым М.В.Волькенштейном: «Живые тела, существующие на Земле, представляют собой открытые саморегулирующиеся и самовоспроизводящиеся системы, построенные из биополимеров – белков и нуклеиновых кислот».
2.3 Изменчивость – это свойство, противоположное наследственности. Оно связано с приобретением организмами новых признаков и свойств. В основе изменчивости лежат мутации – нарушение процесса самовоспроизведения ДНК. Изменчивость создает материал для естественного отбора.
2.4 Свойством живых организмов является способность к историческому развитию и изменению от простого к сложному. Этот процесс называется эволюцией. В результате эволюции возникло все многообразие живых организмов, приспособленных к определенным условиям существования.
Некоторые исследователи к основным свойствам живых организмов относят также: а)единство химического состава (98% - С, N, О, Н);
б)сложность и высокую степень организации, т.е. усложненное внутреннее строение, но в настоящее время обнаружены живые организмы, образованные одной молекулой – прионы – белки.
Эмбриональное развитие
Оно протекает как непрерывный процесс изменения зародыша, однако для удобства изучения его подразделяют на периоды:
- одноклеточный;
- дробления и образования гаструлы;
- период образования тканей и органов, заканчивающийся формированием организма, готового к рождению.
Оплодотворение. Его результатом является возникновение одноклеточного зародыша. Важнейшими при оплодотворении являются 2 момента. Первый – слияние гаплоидных ядер мужской и женской гамет с образованием диплоидной зиготы. Второй – активизация яйцеклетки и побуждение ее к развитию.
Дробление. По своей сути это ряд непрерывно следующих одно за другим митотических делений яйцеклетки.
П.И. Балинский так характеризует этот процесс.
1 Одна клетка – оплодотворенное яйцо – посредством ряда митотических делений превращается в многоклеточный комплекс.
2 Рост отсутствует.
3 Общая форма зародыша в процессе дробления не меняется, но образуется внутренняя (первичная) полость тела.
4 В процессе дробления имеет место превращение веществ, запасенных в цитоплазме, в субстанции ядра.
5 Взаимное расположение частей цитоплазмы яйца в процессе дробления по большей части не меняется, и они остаются на своих местах.
6 Ядерно – плазменное отношение, низкое в начале дробления, достигает в конце уровня, характерного для обычных соматических клеток.
Соответственно типу яйцеклеток, дробление происходит различно.
У низших хордовых, мелких примитивных животных яйцеклетки мелкие, с небольшим количеством желтка. Дробление у них полное, равномерное. У низших позвоночных, более сложно устроенных, (рыбы, земноводные) яйцеклетки крупнее, содержат неравномерно распределенный запас желтка, поэтому дробление хоть и полное, но не равномерное. У высших позвоночных (пресмыкающиеся, птицы) яйцеклетки крупные, богаты желтком. Дробление у таких животных неполное, поверхностное. Оно происходит только в зародышевом диске, т.е. участке, свободном от желтка.
Млекопитающие и человек занимают особое место среди высших позвоночных. Развитие зародыша происходит внутри организма матери, обеспечивающего питание зародыша. В связи с этим яйцеклетки мелкие, желтка – небольшое количество. Для них типично полное равномерное дробление.
Гаструляция. С завершением процесса дробления формированием зародыша (бластулы) начинается период гаструляции. Характерная черта этого периода – массовое, упорядоченное по направлению и последовательности, перемещение клеток. В результате зародыш из однослойного становится многослойным. Образуются зародышевые листки. Это пласты клеток, занимающих определенное место друг относительно друга. Из них в последующем закономерно развиваются определенные органы и части тела. Наружный зародышевый листок называют эктодермой, внутренний – энтодермой, средний - мезодермой. Зародыш в периоде гаструляции называется гаструлой.
Органогенез. Это процесс образования органов. Параллельно развиваются и ткани. Этот процесс называют гистогенезом. Сложные пространственные преобразования клеток и частей зародыша получили название морфогенез.
Каждый зачаток органа, а затем и орган, развиваются из клеток определенных зародышевых листков.
Из эктодермы образуются вся нервная система, определенные части органов чувств, например, роговица глаза, кожный эпителий и его производные (молочные, потовые и сальные железы, волосы, ногти, эмаль зубов), эпителий полости рта и прямой кишки.
Из мезодермы развиваются скелет позвоночника, черепа и конечностей, поперечно – полосатая скелетная мускулатура, гладкая мускулатура внутренних органов, кровеносная система и кровь, выделительная система, половые железы, за исключением самих половых клеток, вся соединительная ткань.
Энтодерма дает начало эпителию пищеварительной и дыхательной систем, печени, поджелудочной железе, щитовидной и паращитовидной железе.
Зародышевые оболочки . У всех высших позвоночных, начиная с пресмыкающихся, образуются особые временные органы, предназначенные для обеспечения важнейших функций зародыша. Таких зародышевых оболочек 4. Они образуются после гаструляции из зародышевых листков.
Амнион, или водная оболочка, окружает непосредственно зародыш. Внутри него содержится жидкость, которая предохраняет зародыш от высыхания и механических повреждений. Амнион человеческого зародыша называется плодным пузырем и также содержит жидкость, которая отходит при его разрыве во время родов.
Хорион. Он способствует дыханию и питанию зародыша. У птиц хорион ближе всего прилежит к скорлупе, а у млекопитающих входит в состав плаценты и прилежит к стенке матки матери.
Аллантоис, мочевой мешок, служит у птиц для выделения жидких азотсодержащих продуктов обмена и для дыхания; (у млекопитающих наряду с хорионом входит в состав плаценты).
Желточный мешок. У птиц служит для питания, дыхания и кроветворения. У млекопитающих он не имеет функции питания в связи с переходом к чрезплацентарному способу поступления питательных веществ, но служит источником половых клеток и клеток крови.
Зародышевые оболочки прекращают свое существование после рождения, выходят наружу вместе со слизистой оболочкой матки.
Близнецы. У человека и некоторых млекопитающих, таких как лошади и коровы, обычно рождается один детеныш. Если же их больше, то говорят о близнецах.
Различают 2 вида близнецов: однояйцовые (ОБ) и разнояйцовые (РБ). Основной механизм возникновения ОБ – разделение зародыша на 2 части или более в периоде дробления эмбриогенеза. Причины этого пока не ясны. Частота рождения РБ связана с индивидуальными особенностями родителей. Описаны семьи, в которых близнецы рождались в нескольких поколениях. Зародыши на ранних стадиях способны как бы исправлять ошибки, и из каждой отделившейся части развивается вполне нормальный организм. Способность ранних стадий зародыша к развитию в нормальное целое после изменения их клеточного состава называется эмбриональной регуляцией и в полной мере проявляется в развитии ОБ. РБ развиваются из двух разных яйцеклеток, оплодотворенных разными сперматозоидами.
Иногда происходит неполное разделение клеточной массы бластомеров на ранних стадиях эмбриогенеза. В таких случаях появляются неразделившиеся двойни вследствие нарушения развития зародыша на 1-й неделе после оплодотворения. Они всегда однояйцовые. Неразделившиеся близнецы могут быть симметричными и соединяться разными отделами тела, например, в области груди, живота, крестца. Чаще всего они нежизнеспособны и погибают в периоде внутриутробного развития. Однако возможны и исключения. Близнецы, родившиеся в Сиаме (Таиланд), и прожившие 63 года, дали общее название таким близнецам (сиамские близнецы).
Врожденные пороки и критические периоды развития человека.
Кроме двойниковых уродств, встречаются новорожденные с различными иными отклонениями. Врожденным пороком развития называется стойкое изменение строения органа, приводящее к расстройству его функции. Количество форм пороков исчисляется тысячами. Различают пороки развития лица и шеи, нервной системы, глаз, опорно-двигательного аппарата, сердечно-сосудистой, дыхательной, пищеварительной, выделительной и половой систем.
Нарушения эмбриогенеза происходят на клеточном, тканевом, органном и организменном уровнях. Отсюда следует, что основными механизмами врожденных пороков являются нарушения наиболее важных процессов, таких как размножение, дифференцировка, перемещение и гибель клеток, рост и взаимодействие развивающихся тканей и органов. Помимо наследственных пороков есть и пороки, возникшие в результате действия экзогенных факторов. К таким факторам относятся:
- физические (проникающая радиация);
- химические (медикаменты, ядовитые вещества, гипоксия зародыша, эндокринные заболевания матери);
- биологические агенты (вирусы).
Критическими периодами являются такие, во время которых чувствительность зародыша к действию внешних факторов особенно велика. У человека 1-й период приходится на конец 1-й начало 2-й недели беременности; 2-й с 3 по 6 неделю, хотя нет таких стадий, в которых эмбрион был бы стоек ко всем повреждающим воздействиям.
Постэмбриональное развитие
Постэмбриональное развитие начинается с момента выхода развивающегося организма из оболочек яйца или организма матери. Главной особенностью его является то, что он вступает в непосредственный контакт с внешней средой. Он начинает дышать, питаться, реагировать на различные воздействия. После рождения развитие организма продолжается в виде изменений в росте, дальнейшей специализации клеток и тканей, регенерации и старении.
РАЗДЕЛ 2 МИКРОБИОЛОГИЯ
Лекция № 4 Введение
Дрожжи Общая характеристика
Дрожжи являются одноклеточными неподвижными микроорганизмами, широко распространенными в природе; они встречаются в почве, на листьях, стеблях и плодах растений, в разнообразных пищевых субстратах растительного и животного происхождения.
Широкое использование дрожжей в промышленности основано на их способности вызывать спиртовое брожение.
Форма и строение дрожжевой клетки. Форма клеток дрожжей чаще округлая, овально-яйцевидная или эллиптическая, реже цилиндрическая и лимоновидная. Встречаются дрожжи особой формы - серповидные, игловидные, стреловидные, треугольные. Размеры дрожжевых клеток обычно не превышают 10—15 мкм.
Форма и размеры дрожжей могут заметно изменяться в зависимости от условий развития, а также возраста клеток.
Строение клетки дрожжей сходно со строением клетки грибов. Дрожжи обладают всеми основными структурами, характерными для эукариотного типа клетки (ядро, отграниченное от цитоплазмы, эндоплазматическая сеть, аппарат Гольджи, лизосомы, митохондрии, рибосомы, вакуоли). В качестве запасных питательных веществ в клетках обнаруживаются капельки жира, гранулы гликогена, волютина.
Клеточная стенка (оболочка) дрожжей слоиста, в состав ее у большинства дрожжей входят в основном (до 60—70% сухой массы) гемицеллюлозы; в небольших количествах — белки, липиды, хитин. У некоторых дрожжей оболочка может в той или иной степени ослизняться, вследствие чего клетки склеиваются друг с другом и при развитии в жидких средах образуют оседающие на дно сосуда хлопья. Такие дрожжи называют хлопьевидными, в отличие от пылевидных, клеточные стенки которых не ослизняются; пылевидные дрожжи в жидкости находятся во взвешенном состоянии.
Размножение дрожжей. Наиболее характерным и широко распространенным у дрожжей вегетативным способом размножения является почкование, лишь немногие дрожжи размножаются делением.
У дрожжей известны следующие типы почкования: мультилатеральное, биополярное, униполярное и равномерное.
Процесс почкования заключается в том, что на клетке появляется бугорок (иногда их несколько), который постепенно увеличивается. Этот бугорок называют почкой. Почкованию предшествует разделение ядра на две части, и оно вместе с частью цитоплазмы и другими клеточными элементами переходит в формирующуюся молодую клетку. По мере роста почки в месте соединения ее с материнской клеткой образуется перетяжка, отграничивающая молодую дочернюю клетку, которая затем либо отшнуровывается (отделяется) от материнской клетки, либо остается при ней. В месте отделения дочерней клетки остается рубец. При благоприятных условиях этот процесс длится около 2 ч.
Почкующиеся клетки обычно образуют не одну, а несколько почек. Вместе с этим может начаться почкование и молодых клеток. Так постепенно образуются скопления из многих объединенных между собой клеток, называемые сростками почкования.
В некоторых случаях, особенно на поверхности жидких сред, такие сростки почкования образуют тонкую пленку, легко разрушающуюся при взбалтывании жидкости. Существуют дрожжи, которые образуют более или менее толстые морщинистые пленки, прочно удерживающиеся при взбалтывании. Такие пленчатые дрожжи нередко вызывают порчу соленых и квашеных овощей, вина, пива.
Помимо почкования, многие дрожжи размножаются с помощью спор. Споры у дрожжей могут образовываться бесполым и половым путями. В первом случае ядро клетки делится на столько частей, сколько образуется спор у данного вида дрожжей, после чего постепенно в клетке (как в сумке) образуются аскоспоры. Образованию спор половым путем предшествует слияние (копуляция) клеток. У некоторых дрожжей копулируют прорастающие споры. Число спор в клетке разных видов дрожжей различно. Их может быть две, четыре, а иногда восемь и даже двенадцать.
Споры большинства дрожжей округлые или овальные, но у некоторых игловидные, шляповидные. На поверхности многих спор имеются различные образования типа выростов, бородавок, ободков. Споры дрожжей более устойчивы к неблагоприятным воздействиям, чем вегетативные клетки, но менее стойки, чем бактериальные споры. В благоприятных условиях споры прорастают в клетки.
У многих так называемых культурных дрожжей, т. е. культивируемых человеком для производственно-хозяйственных целей, способность к спорообразованию в значительной степени ослаблена, а иногда полностью утрачена.
Основы систематики дрожжей. Дрожжи, как указывалось выше, относятся к классу сумчатых грибов (Ascomycеtes), к подклассу голосумчатых, не образующих мицелия. Разделение голосумчатых грибов на порядки, семейства, роды основано на особенностях их размножения, морфологических, физиологических и биохимических признаках.
Наибольший интерес представляет род сахаромицес (Saccharomyces), который объединяет как природные виды, так и культурные, применяемые в промышленности. Отдельные их виды различаются способностью сбраживать те или иные сахара, интенсивностью брожения, количеством образуемого спирта, оптимальными температурами почкования и образования спор и т. д.
В промышленности наиболее широко используют дрожжи сахаромицес церевизия. В настоящее время в различных странах мира их вырабатывают более 2 блн т. Коммерческими продуктами являются прессованные и высушенные различными способами дрожжи, а также пищевые дрожжи, характеризующиеся полностью инактивированными ферментными системами. Пищевые дрожжи используют как добавки к продуктам питания, а не как биологические катализаторы.
Сахаромицес церевизия (Sacch. cerevisiae) — дрожжи округлой или овальной формы. Применяют их в производстве этилового спирта, пивоварении, квасоварении и хлебопечении. Каждое производство применяет свои специфические расы (разновидности) данного вида дрожжей.
Сахаромицес вини (Sacch. vini) — дрожжи эллиптической формы. Их используют преимущественно в виноделии. Этот вид дрожжей также представлен многими расами.
Эти и некоторые другие виды рода Saccharomyces при спонтанном (самопроизвольном) развитии в содержащих сахар пищевых продуктах вызывают их порчу — забраживание, прокисание.
Помимо спорообразующих, существуют дрожжи, не образующие спор, — аспорогенные. Нередко их называют дрожжеподобными или несовершенными дрожжевыми организмами и относят к несовершенным грибам.
Из аспорогенных дрожжей наибольшее значение имеют роды кандида (Candida) и торулопсис (Torulopsis). Многочисленные представители их широко распространены в природе, большинство не способно к спиртовому брожению, многие вызывают порчу пищевых продуктов.
Торулопсис имеют клетки округлой или овальной формы. Многие из них способны вызывать лишь слабое спиртовое брожение. Отдельные виды используют в производстве кумыса и кефира.
Кандида — дрожжи, клетки которых имеют вытянутую форму, способны к образованию примитивного мицелия (псевдомицелий). Многие из них не способны к спиртовому брожению. Некоторые виды (например, Кандида утилис), окисляющие сахар и этиловый спирт в органические кислоты или в углекислый газ и воду, являются вредителями в производствах вин, пива, пекарских дрожжей. Эти дрожжи вызывают порчу квашеных овощей, безалкогольных напитков и многих других продуктов.
Имеются виды, вызывающие заболевания — кандидозы у людей, при которых поражаются слизистые оболочки рта и других органов.
Среди аспорогенных дрожжей имеются окрашенные в желтый, розовый, красный цвета, что обусловлено наличием в клетках пигментов — каротиноидов. В настоящее время некоторые из этих дрожжей (виды рода родоторула —Rhodotorula) используют для получения кормовых белково-каротиноидных препаратов, которые являются источником витамина А для животных.
В последнее время дрожжи Кандида утилис широко применяют для получения белка, аминокислот, витаминов и ферментов. Например, синтезирует белок при выращивании ее на отходах бумажной промышленности, а Сахаромицес липолитика— на алканах нефти; Сахаромицес церевизия используют для получения фермента инвертазы.
В МИНХе им. Г. В. Плеханова А. А. Кудряшовой с группой сотрудников получен препарат из дрожжей "Александрина", предназначенный для использования в лечебно-профилактических целях. Он представляет собой полностью растворяющуюся в воде смесь аминокислот, витаминов и минеральных элементов. В пищевой промышленности он может применяться в качестве высокоэффективной и полифункциональной натуральной пищевой добавки, не оказывающей негативного влияния на органолептические свойства продуктов питания. Добавление этого препарата к пищевым продуктам способствует повышению их физиологической ценности, появлению новых положительных признаков и свойств или усилению им присущих (повышение качества, усиление пеностойкости напитков и желеобразующей способности кондитерских изделий, замедление черствения хлебобулочных изделий, сохранение формы мучных изделий при варке, снижение калорийности сахаристых изделий и др.). Отходы, образующиеся при производстве этого препарата, могут использоваться как в пищевой промышленности, так и в области животноводства, растениеводства, при выращивании лабораторных и промышленных микроорганизмов, пушных зверей, рыбы, шляпочных грибов, цветов и др.
Новая натуральная биопродукция позволяет получать экологически безопасные продукты питания, эффективные корма для животных и питательные среды для микроорганизмов, новые медицинские препараты и парфюмерно-косметические изделия высокой биологической активности. Разрешение на ее промышленное производство и реализацию для населения получены в Минздраве РФ и в Госстандарте.
3.3 Вирусы и фаги. Вирусы (от лат. Virus - яд) - это особая группа микроорганизмов меньших размеров и более простой организации, чем бактерии. Вирусы не имеют клеточной структуры, величина их измеряется нанометрами. Вирусы открыты русским ботаником Д. И. Ивановским в 1892 г. при изучении мозаичной болезни листьев табака, которая причиняла большой ущерб табачным плантациям Крыма. Открытие Д. И. Ивановского заложило основу новой науки - вирусологии.
Вирусы — внутриклеточные паразиты, вызывающие многие болезни человека (оспу, грипп, бешенство, корь, полиомиелит и др.), животных (ящур, чуму крупного рогатого скота) и растений ("мозаики" и другого вида заболевания полевых и огородных культур).
Вирусы разнообразны по форме, размерам и химическому составу. Большинство из них имеет палочковидную или сферическую форму. Некоторые вирусы состоят только из белка и одной нуклеиновой кислоты - ДНК или РНК, другие содержат еще и липиды, полисахариды. Вирусная частица называется вирионом. Нуклеиновая кислота (в виде спирали) находится внутри вириона, снаружи он покрыт белковой оболочкой (капсидом), состоящей из отдельных морфологических субъединиц (капсомеров). Вирусы выращивают на живых клетках или культуре тканей, так как на искусственных питательных средах они, как правило, не развиваются.
Вирусы обладают разной устойчивостью к внешним воздействиям. Многие инактивируются при 60"С до 10 мин, другие выдерживают температуру 90°С до 10 мин. Вирусы довольно легко переносят высушивание и низкие температуры, но мало устойчивы ко многим антисептикам, ультрафиолетовым лучам, радиоактивным излучениям.
Фаги - это вирусы микроорганизмов, вызывающие гибель — распад (лизис) их клеток. Вирусы бактерий называются бактериофагами или просто фагами, актиномицетов — актинофагами, вирусы грибов — микофагами, сине-зеленых водорослей (цианобактерий) - цианофагами.
Впервые лизис сибиреязвенных бактерий наблюдал Н. Ф. Гамалея в 1898 г. Д. Эррель в 1917 г. установил явление лизиса у бактерий дизентерии, им впервые был выделен и описан бактериофаг ("пожиратель") бактерий.
Морфология фага изучена с применением электронного микроскопа. Большинство фагов состоит из головки и отростка. Головка фага может иметь разную форму, чаще всего это многогранник, покрытый белковой оболочкой (капсидом). Внутри капсида расположена нуклеиновая кислота, чаще всего одна — ДНК или РНК. Отросток фага имеет внутренний полый стержень, по каналу которого ДНК фага переходит в клетку хозяина. Стержень снаружи покрыт чехлом, способным к сокращению. Стержень и чехол отростка состоят из белковых субъединиц. У некоторых фагов отросток заканчивается базальной пластинкой, которая имеет выступы (зубцы) и нити.
Фаги могут быть и нитевидной формы, могут состоять из одной головки, а могут быть с аналогами отростка (очень коротким отростком). Некоторые фаги имеют длинные отростки с несокращающимся или сокращающимся чехлом.
Фаги широко распространены в природе. Многие из них обладают специфичностью — могут воздействовать на определенный вид или группу родственных видов микроорганизмов.
Взаимодействие фага с микробной клеткой происходит в несколько фаз. Сначала фаг адсорбируется восприимчивой клеткой, затем под действием фермента фага (сходного с лизоцимом) в стенке микробной клетки образуется отверстие, через которое в клетку проникает только нуклеиновая кислота; пустая белковая оболочка головки и отростка остается снаружи клетки, а затем разрушается.
Под влиянием попавшей в клетку нуклеиновой кислоты фага перестраиваются все обменные процессы микробной клетки на синтез фаговых частиц: синтезируются фаговая нуклеиновая кислота и белковые субъединицы оболочек. Вначале формируются раздельно головки и отростки, которые затем объединяются в зрелые фаговые частицы. Через определенное время клетка хозяина погибает, разрушается и фаги выходят наружу.
Явление фаголизиса (растворение культур микроорганизмов) наблюдается на производствах, связанных с использованием микроорганизмов. Развитие фагов в культурах промышленных микроорганизмов приводит к тому, что клетки культуры лизируются, не успев синтезировать необходимые вещества. Это наносит предприятиям большой экономический ущерб. Так нередко лизируются молочно-кислые бактерии, входящие в состав заквасок для кисломолочных продуктов. Такие закваски не пригодны для употребления.
Бактериофаги , лизирующие зараженные ими бактерии, называют вирулентными. Некоторые фаги, однако, инфицируют бактерии, но не вызывают их лизиса; такие фаги называются умеренными. В клетке-бактерии - хозяина они не размножаются, но при делении бактерии передаются дочерним клеткам.
Фаги применяются в медицине для лечения и профилактики некоторых заболеваний, например дизентерии, холеры. Фаги исключительно удобны как модели для решения вопросов общебиологических, молекулярной биологии, генетики, медицины.
Лекция № 7. Физиология микроорганизмов. Химический состав микроорганизмов. Питание микробной клетки. Гетеротрофный и автотрофный типы питания. Конструктивный и энергетический обмен. Рост и размножение микроорганизмов.
Физиология микроорганизмов.
Изучает процессы их роста, развития, питания, способы получения энергии для осуществления этих процессов, их взаимодействия с окружающей средой. Знание физиологических процессов микроорганизмов создает научную основу для проведения культивирования (выращивания) и идентификации (распознавания) видов микробов, а также получения биологических и лечебных препаратов (заквасок, витаминов, ферментов, аминокислот, антибиотиков, вакцин и др.).
Понятие об обмене веществ. Основу жизнедеятельности микроорганизмов, как и всех живых существ, составляет обмен веществ (метаболизм) с окружающей средой. Термин метаболизм объединяет два взаимосвязанных, но противоположных процесса - анаболизм и катаболизм.
Анаболизм (питание; ассимиляция; конструктивный или строительный обмен; обмен веществ) сводится к усвоению, т.е. использованию микробами питательных веществ, поступивших из внешней среды, для биосинтеза компонентов собственного тела. Это достигается чаще восстановительными эндотермическими реакциями, для течения которых требуется энергия.
Катаболизм (дыхание, диссимиляция, биологическое окисление) характеризуется расщеплением (окислением) сложных органических веществ до более простых продуктов с освобождением заключенных в них энергии. Эта энергия используется микроорганизмами для синтеза веществ данной клетки.
Метаболизм у микроорганизмов характеризуется интенсивным потреблением питательных веществ. Например, при благоприятных условиях в течение суток одна клетки бактерий усваивает веществ в 30-40 раз больше величины своей массы, соответственно высока и скорость прироста биомассы микроорганизмов. Основная часть пищи расходуется микроорганизмами в энергетическом обмене, при котором в среду выделяется большое количество продуктов обмена: кислот, спиртов, диоксид углерода, водород и др. Эта особенность микроорганизмов широко используется в практике переработки растительного, животного пищевого и непищевого сырья и обуславливает порчу пищевого сырья.
2 Химический состав микроорганизмов. Состав веществ тела микроорганизмов мало отличается от состава тела растений и животных.
Для определения потребностей микроорганизмов в питательных веществах необходимо знать их химический состав. Элементарный состав клеток микроорганизмов довольно разнообразен и представлен в процентах от сухого вещества клетки: углерод-50, кислород-20, азот-14, водород-8, фосфор-3, сера, калий , натрий- по 1, кальций, магний, хлор- по 0,5, железо-0,2, все остальные по 0,3. Эти элементы играют различную физиологическую роль. Так, углерод, кислород, азот и водород входят в состав всех без исключения живых организмов, их называют органогенами. Эти элементы составляют основу органических веществ; водород и кислород входят в состав воды; кислород необходим для дыхания аэробным микроорганизмам. Важную физиологическую функцию выполняют также фосфор и сера. Фосфор входит в состав важных органических соединений клетки - фосфолипидов, АТФ и др. Сера необходима для серосодержащих аминокислот (цистина, цистеина, гомоцистеина, метионина), без которых невозможен синтез белков.
Микробная клетка состоит из воды и сухих веществ. Количество воды для большинства микробов колеблется от 75 до 85% и находится в клетке в свободном и связанном состояниях, что имеет важное значение в жизни микроорганизмов, так как все вещества поступают в клетку только с водой и с ней же удаляются продукты обмена из клетки. Свободная вода служит дисперсной средой для коллоидов и растворителем различных органических и минеральных соединений. Связанная вода является структурным элементом цитоплазмы и не может быть растворителем. Содержание воды в клетке изменяется в зависимости от условий внешней среды, физиологического состояния клетки, ее возраста и т.п. В спорах бактерий и грибов значительно меньше воды, чем в вегетативных клетках, за счет низкого содержания в них свободной воды. Потеря свободной воды влечет за собой высыхание клетки и изменения в обмене веществ. С потерей связанной воды нарушаются клеточные структуры и наступает гибель клетки.
Сухое вещество клеток микроорганизмов не превышает 15-25% и состоит преимущественно (до85-95%) из органических соединений- белков, углеводов, нуклеиновых кислот, липидов и других соединений.
Белковые вещества являются основными компонентами клетки. Содержание их зависит от вида микроорганизмов, условий выращивания и возраста и составляет в среднем от 40 до 60%. По аминокислотному составу белки микроорганизмов сходны с белками других организмов. Некоторые белки (ферменты) выполняют каталитические функции: осуществляют различные биохимические реакции, постоянно протекающие в микробной клетке).
Многие микроорганизмы могут накапливать большое количество белков в составе своих клеток и их можно рассматривать в качестве продуцентов пищевого и кормового белка. Рентабельность промышленного производства таких «белковых продуктов» определяется быстротой накопления биомассы микроорганизмов и использованием для их выращивания дешевого недефицитного сырья (отходов различных производств).
Углеводы составляют 15-20% сухого вещества и содержатся в микробных клетках в основном в виде полисахаридов. Углеводы входят в состав капсул. Клеточных мембран и цитоплазмы, а также являются запасными веществами в виде включений гранулезы и гликогена.
Нуклеиновые кислоты содержатся в клетках в виде рибонуклеиновой (РНК) и дезоксирибонуклеиновой (ДНК) кислот. ДНК сосредоточена главным образом в ядре эукариотных клеток и в нуклеодах прокариотных (бактериальных клеток). В молекуле ДНК закодирована вся наследственная информация клетки, «записаны» все особенности будущего организма, выработанные в процессе длительной эволюции и свойственные данному виду. РНК преимущественно сосредоточена в цитоплазме и рибосомах.
Липиды составляют 3-10% сухого вещества, входят в состав клеточных оболочек и надежно защищают клетку от воздействий окружающей среды, а также откладываются в виде запасных гранул. Часть липидов связана с другими веществами клетки, образуя сложные комплексы (эфиры сложных кислот и углеводов, воски и фосфолипиды). Пигменты и красящие вещества обуславливают окраску микроорганизмов. Фотосинтезирующие бактерии содержат особые пигменты типа хлорофилла растений - бактериохлорофилл. Фототрофные бактерии и некоторые дрожжи образуют пигменты - каротиноиды, которые участвуют, как и бактериохлорофилл, в ассимиляции углекислого газа. У некоторых грибов (мицелиальных и дрожжевых) в значительных количествах образуются желто-розовые и оранжевые каротиноиды, которые являются провитаминами витамина А.
Минеральные вещества составляют 5-15% сухого вещества клетки и представлены сульфатами, фосфатами, карбонатами, хлоридами. Фосфаты могут быть в свободном виде и входить в состав различных соединений (нуклеиновых кислот, АДФ, АТФ). Минеральные соединения играют важную роль в регулировании внутриклеточного давления и коллоидного состояния цитоплазмы. Они влияют на скорость и направление биохимических реакций, являются стимуляторами роста, активаторами ферментов.
3 Питание микробной клетки. Анаболизм микроорганизмов. Питание-это процесс усвоения микробной клеткой питательных веществ, поступающих из окружающей среды, в результате которого они превращаются в составные части биологических структур клетки или откладываются в ней в виде запасов. Большинство микроорганизмов, также как и растения, обладают голофитным способом питания, или внеклеточным (внешним) пищеварением, которое происходит в окружающей среде (субстрате) под действием экзоферментов микроорганизмов.
Существует также голозойный способ питания (внутриклеточное пищеварение), которое происходит под действием эндоферментов. Оно присуще простейшим и некоторым низкоорганизованным организмам и характеризуется заглатыванием (обволакиванием) плотных частиц пищи, перевариванием и превращением их в растворимые соединения (эндоцитоз).
Возможность проникновения веществ извне в клетку обусловлено многими факторами: величиной и структурой их молекул; способностью растворяться в компонентах цитоплазматической мембраны; концентрацией веществ в клетке и в среде. Имеют значение также и свойства клеточной стенки и цитоплазматической мембраны, являющихся барьерами, через которые должны проникнуть в клетку питательные вещества; имеет значение электрический заряд поверхности клетки и др.
Вещества питательной среды могут поступать в клетку в растворенном состоянии. Нерастворимые сложные органические соединения должны подвергнуться расщеплению на более простые вне клетки, что происходит с помощью экзоферментов микроорганизмов.
Клеточная стенка проницаема и задерживает лишь макромолекулы. Цитоплазматическая мембрана обладает полупроницаемостью и служит осмотическим барьером, так как проницаемость ее для различных веществ неодинакова. Известно несколько путей проникновения питательных веществ в клетку.
Пассивная диффузия подчиняется законам осмоса. При осмотическом проникновении веществ через полупроницаемую мембрану движущей силой является разность осмотических давлений (концентраций веществ) в растворах по обе стороны мембраны, т.е. между средой и клеткой. При этом концентрация растворенных солей внутри клетки несколько выше (по сравнению с субстратом), а так как вода по закону осмоса стремится в сторону противоположной концентрации, то она поступает в клетку, увлекая с собой питательные вещества. Такой пассивный перенос веществ (по градиенту концентрации) протекает до выравнивания концентраций и не требует затраты энергии. При этом внутреннее напряжение клетки (так называемый тургор) является одним из основных условий, обеспечивающих нормальное поступление в нее питательных веществ. Для большинства микроорганизмов тургор наиболее выражен при 0,85%-ной концентрации солей в окружающей среде. Эта концентрация называется изотонической.
При гипертонической концентрации, т.е. при повышении ее до 2-3%, наступает плазмолиз (обезвоживание) - сжатие, сморщивание цитоплазмы и отслаивание ее от клеточной стенки. При помещении микроорганизмов в гипотонический раствор (например, дистиллированная вода) – вода прямым потоком поступает внутрь клетки. Объем клетки при этом увеличивается, происходит набухание и разрушение оболочки клеток. Это явление получило название плазмоптиза.
Облегченная диффузия. Скорость транспорта веществ в клетку в условиях повышения концентрации субстрата возрастает до определенного предела. При облегченной диффузии кроме градиента концентрации функционируют электрические переносчики, находящиеся в мембране: субстрат соединяется с протоном и белком - переносчиком и по электрическому градиенту диффундирует в клетку. Переносчики являются специфичными по отношению к субстрату. Так, дрожжевые клетки поглощают сахара путем облегченной диффузии, а у анаэробов этим способом происходит поглощение некоторых соединений и выделение продуктов обмена.
Активный транспорт. Поступление питательных веществ осуществляется против градиента концентраций с затратами энергии со стороны клетки и при участии специфических белков-переносчиков (пермеаз), локализованных в цитоплазматической мембране. Пермеазы сходны с ферментами и обладают субстратной специфичностью - каждая транспортирует определенное вещество. На внешней стороне цитоплазматической мембране пермеаза адсорбирует вещество, вступает с ним во временную связь и диффундирует комплексно через мембрану, отдавая на внутренней стороне ее в цитоплазму. Выход растворенных веществ осуществляется как при участии пермеаз - путем облегченной диффузии, так, видимо, и путем пассивной диффузии.
4 Конструктивный обмен. Пища должна содержать такие вещества, которые удовлетворяли бы потребность микроорганизмов в химических элементах и энергии, необходимых для синтеза веществ и структур клетки. В зависимости от того, какие химические элементы поступают из веществ питательной среды, их называют источниками углерода, азота, фосфора и др.
Кислород и водород микроорганизмы получают из воды, содержатся они также во многих используемых органических соединениях.
Потребности микроорганизмов в отношении источников углерода и азота весьма разнообразны.
Источники углерода. В зависимости от используемого в конструктивном обмене источника углерода микроорганизмы делят на две группы: автотрофы (питающийся самостоятельно) и гетеротрофы (питающийся другими).
Автотрофы усваивают углерод из диоксида углерода воздуха и в зависимости от вида используемой энергии для фиксации СО2 их соответственно называют фото- и хемосинтезирующими.
Фотосинтезирующие используют энергию солнечных лучей. Они напоминают зеленые растения, образующие в процессе фотосинтеза углеводы из СО2 и Н2О. К этой группе относят цветные бактерии, имеющие в цитоплазме своих клеток пигменты типа хлорофилла, как, например, пурпурные серные бактерии, сине-зеленые водоросли и др.
Хемоситезирующие используют энергию химических реакций окисления минеральных (неорганических) веществ. К ним относят нитрифицирующие бактерии, бесцветные серобактерии, железобактерии, нитчатые, тионовые, водородные бактерии и др.
Гетеротрофы - наиболее многочисленная группа микроорганизмов, которые могут использовать углерод только из готовых органических субстратов животного и растительного происхождения. Микроорганизмы называют сапрофитами (метатрофами), если они используют мертвый органический субстрат. Они разлагают различные органические вещества в природе, вызывают порчу пищевых продуктов или используются в процессах переработки растительного и животного сырья. Многие сапрофиты всеядны, т.е. способны использовать разнообразные органические соединения. Существуют и такие, которые нуждаются в определенных соединениях (например, микробы, использующие метан, парафины и др.) Сапрофиты, наряду с органическими соединениями, используют в небольших количествах и СО2, который служит дополнительным источником углерода для биосинтеза веществ клетки. Вторая подгруппа гетеротрофов - паразиты (паротрофы) питаются живыми органическими субстратами. Сюда относятся преимущественно болезнетворные микроорганизмы, т.е. паразитирующие в живых организмах. Абсолютными (внутриклеточными) паразитами являются вирусы и риккетсии, которые развиваются в живых клетках человека, животных, растений и микроорганизмов.
В отличие от вирусов и риккетсий многие патогенные микроорганизмы - паразиты могут размножаться на искусственных питательных средах, т.е. по типу питания занимают промежуточное положение между паразитами и сапрофитами (патогенные стафилококки, стрептококки, возбудители бруцеллеза, туберкулеза, сибирской язвы и др. инфекционных болезней).
Источники азота. В зависимости от источника азота все микроорганизмы можно разделить также на две группы: аминоавтотрофы и аминогетеротрофы.
Аминоавтотрофы усваивают азот из неорганических источников. Они представлены двумя подгруппами: азотфиксирующие и нитритно-нитратные микроорганизмы. Азотфиксирующая подгруппа способна усваивать молекулярный азот воздуха (актиномицеты, азотфиксирующие - свободноживущие и симбиотические). Нитритно - нитратные микроорганизмы окисляют аммиак до солей азотистой и азотной кислот и усваивают эти окисленные формы азота.
Аминогетеротрофы используют органические источники азота. К ним относятся дезаминирующие, пептонные, протеолитические и паротрофные микроорганизмы. Дезаминирующие микроорганизмы могут усваивать только аминокислоты (некоторые патогенные бактерии). Пептонные бактерии потребляют только органические соединения типа пептонов, так как не способны расщеплять цельную белковую молекулу (молочнокислые, пропионовокислые бактерии, энтерококки, микрококки и кишечные палочки). Протеолитические микроорганизмы или гнилостные в качестве источника азота используют натуральные белки, которые предварительно разлагаются их экзоферментами (гнилостные бактерии, актиномицеты, плесени). Паротрофные микроорганизмы в качестве источника азота используют белковые вещества живого организма (патогенные).
Установить резкую грань между автотрофами и гетеротрофами не всегда удается. Некоторые патогенные микроорганизмы во внешней среде ведут сапрофитный образ жизни, и наоборот, некоторые сапрофиты в зависимости от состояния микроорганизма могут вызвать заболевания.
Минеральные элементы. Микробная клетка нуждается в минеральных веществах. Потребность в них невелика, но без некоторых элементов невозможны рост и развитие микрооргнизмов. Калий активизирует ферментативные процессы, ускоряет течение физиологических процессов. Магний входит в состав хлорофилла у зеленых и пурпурных серобактерий, активизирует карбоксилазу, пептидазу и другие ферменты. Фосфор входит в состав нуклеиновых кислот, принимает активное участие в процессах дыхания (окисления). Сера – один из компонентов белков , входит в состав некоторых аминокислот. Железо необходимо в малых количествах, входит в состав дыхательных ферментов, ускоряет процессы окисления. Микроэлементы нужны микробной клетке еще в меньших количествах, но их недостаток ведет к нарушению нормального роста и развития. Молибден, бор, марганец, кобальт, медь и др. микроэлементы являются компонентами многих ферментов и витаминов. Для получения этих химических элементов в питательные среды для микроорганизмов вводят минеральные соединения.
Факторы роста. Активаторы биологических процессов по своему действию являются витаминами и витаминоподобными соединениями. Одни микроорганизмы должны получать витамины в готовом виде, а другие синтезируют витамины в количествах, значительно превышающих собственные потребности. На этом основан микробиологический путь получения рибофлавина (витамин В2), каротиноидов (провитамин А), эргостерина (провитамин Д).
Культивирование микробов в условиях лабораторий осуществляется на искусственных питательных средах. Для гетеротрофов среды должны содержать экстракты из продуктов животного и растительного происхождения с добавлением пептона. Пептон - универсальный источник азота, являющийся продуктом неполного расщепления белков посредством фермента пепсина в кислой среде. В отличие от животных многие микробы могут использовать самые различные субстраты в качестве продуктов питания. Они растут на бумаге, дереве, коже, резине и т.д. Одни из них для своей жизнедеятельности используют парафиновые углеводороды, нефть, керосин; а другие - элективные (избирательные, селективные) среды, имеющие определенный состав.
5 Энергетический обмен. Описанные выше процессы конструктивного процесса – синтез веществ клетки из поступивших в нее извне питательных веществ, активный перенос этих веществ через цитоплазматическую мембрану и многие другие процессы жизнедеятельности - протекают с затратой энергии. Источники энергии у микроорганизмов разнообразны.
У фотоавтотрофов источником энергии служит видимый свет. Световая энергия улавливается фотоактивными пигментами клетки в процессе фотосинтеза, трансформируется в химическую энергию и обеспечивает энергетические потребности клетки.
Источником энергии у хемоавтотрофов служит химическая энергия, получаемая при окислении неорганических соединений (аммиак, сероводород и др.).
Хемогетеротрофы получают энергию в процессе окисления органических соединений. Любое природное органическое вещество и многие синтетические могут быть использованы хемогетеротрофами. Но одни способны окислять многие органические вещества, а другие - лишь небольшой набор их. Некоторые микроорганизмы могут проявлять большую специфичность к энергетическому материалу.
Поскольку все микроорганизмы - возбудители порчи пищевых продуктов и используемые при переработке пищевого сырья относятся к хемогетеротрофам, ниже рассматриваются именно их энергодающие процессы. К ним относят дыхание (1) и брожение(2). Суммарные уравнения выглядят следующим образом:
С6Н12О6 +6 О2=6СО2+6Н2О+674 ккал,
С6Н12О6 → 2СН3СН2ОН+2СО2+27 ккал.
Дыхание (биологическое окисление)- сложный процесс окисления различных органических соединений и некоторых минеральных соединений (нитратов и сульфатов). Нитратное дыхание - восстановление нитратов до молекулярного азота - происходят по схеме. Сульфатное дыхание - восстановление сульфатов до сероводорода, сопровождающееся выделением такого же количества энергии:
5С6Н12О6+24КNО3→24КНСО3+18Н2О+12N2+6СО2+270 ккал.
С6Н12O6+3К2SО4 →3К2СО3+3СО2+3Н2О+3Н2S+270 ккал.
В итоге окислительно-восстановительных процессов и брожения образуется тепловая энергия, часть которой используется микробной клеткой, а остальное количество выделяется в окружающую среду. В настоящее время окисление определяют как процесс отнятия водорода (дегидрирование), а восстановление - его присоединение. Эти термины применяют к реакциям, связанным с переносом протонов и электронов, или только электронов. При окислении вещества происходит потеря электронов, а при восстановлении – их присоединение.
Различают два типа биологического окисления: прямое и непрямое. При прямом окислении органические вещества, такие как молекулярный водород, оксид углерода, метан, сера, аммиак, соли азотистой кислоты, железо и др. окисляются атмосферным кислородом с помощью ферментов оксидаз. При прямом окислении неорганических веществ получают энергию автотрофные почвенные бактерии.
При непрямом окислении происходит отщепление водорода от донора и его присоединение к акцептору. Поэтому непрямое окисление называют дегидрированием. Непрямому окислению путем дегидрирования подвергаются органические вещества при помощи дегидрогеназ. Различают аэробное и анаэробное дегидрирование. При аэробном дегидрировании микроорганизмы используют в качестве конечного акцептора водорода атмосферный кислород. Водород отщепляется от донора с помощью фермента дегидрогеназы и передается акцептору не сразу, а проходит ряд промежуточных этапов.
При аэробном дегидрировании происходит полное и неполное окисление. В случае полного окисления конечными продуктами являются вода и диоксид углерода, происходит высвобождение всей энергии. При неполном окислении освобождается лишь часть энергии. Конечными продуктами неполного аэробного окисления сахара могут быть органические кислоты: лимонная, яблочная, щавелевая, янтарная и др., которые образуются плесневыми грибами. Также осуществляется аэробное дыхание уксуснокислыми бактериями:
СН3СН2ОН+О2→СН3СООН+Н2О+80 ккал
При анаэробном дегидрировании микробы используют в качестве акцептора водорода не кислород, а азот, серу, углерод и другие соединения, которые образуются при распаде субстрата (пировиноградной кислоты). При этом водород довольно легко соединяется с азотом, серой, углеродом, которые восстанавливаются до аммиака, сероводорода и метана.
Дегидрирование углеводов называют брожением, оно чаще проходит в анаэробных условиях. Конечными продуктами брожения являются органические кислоты, этиловый и бутиловый спирты, ацетон и другие продукты. Таким образом, прямое окисление и дегидрирование приводят к одному результату - окислению субстрата, т.е. отщеплению от субстрата водорода и присоединению его к акцептору ( восстановление). Перенос электрона всегда сопровождается освобождением энергии, которая немедленно утилизируется клеткой с помощью особых соединений АТФ и АДФ (аденозинтрифосфата и аденозиндифосфата). В них она накапливается в органических фосфатных (макроэргических) связях и расходуется клеткой по мере необходимости.
По типу дыхания микроорганизмы разделяют на четыре основные группы: облигатные аэробы, облигатные и факультативные- анаэробы и микроаэрофиллы.
Облигатные (строгие) аэробы растут при свободном доступе кислорода воздуха, имеют ферменты, обеспечивающие передачу водорода от донора электронов (субстрата) конечному акцептору - кислороду воздуха. Размножаются при наличии в атмосфере до 21% кислорода, на питательных средах растут на верхних слоях (уксуснокислые бактерии, возбудитель туберкулеза, пигментные гнилостные бактерии, многие плесени и др. микроорганизмы).
Облигатные анаэробы способны к размножению только в атмосфере, свободной от кислорода, или при его содержании не более 5%. У этих микроорганизмов конечным акцептором водорода является субстрат (азотсодержащие вещества, углеводы и др.). Эти микробы растут на дне пробирке под значительным слоем питательной среды. В эту группу входят маслянокислые и пропионовокислые бактерии, гнилостные клостридии, возбудитель ботулизма, бифидобактерии и др. Для некоторых строгих анаэробов кислород является ядом.
Факультативные анаэробы развиваются как при доступе кислорода, так и в его отсутствии. Они имеют набор ферментов, обеспечивающих аэробный и анаэробный тип биологического окисления (дыхания). Развиваются по всей толщине питательной среды. Это многочисленная группа микроорганизмов, к которым относятся молочнокислые бактерии, стафилококки, бактерии группы кишечной палочки, гнилостные бактерии рода Протеус.
Микроаэрофиллы нуждаются в значительно меньшем количестве кислорода, чем аэробы. Они развиваются при концентрации кислорода в окружающей среде не более 10%, т.е. у них преобладает аэробный тип дыхания (актиномицеты, лептоспиры, возбудители бруцеллеза, некоторые плесневые грибы).
Классификация по типам питания. При классификации по типам питания на первое место ставится вид используемой энергии, в соответствии с этим микроорганизмы делят на фототрофы и хемотрофы. Каждую из этих групп в зависимости от окисляемого вещества в свою очередь делят на литотрофы( лито-минерал, камень) и органотротрофы:
1.Фототрофы 2. Хемотрофы
1.1. Фотолитотрофы 2.1. Хемолитотрофы
1.2. Фотоорганотрофы 2.2. Хемоорганотрофы.
Физические факторы.
Температура. Температура внешней среды является мощным фактором воздействия на организмы, который определяет не только интенсивность их развития, но и вообще возможность развития. Принято различать три основные температурные точки, имеющие значение для развития микробов: температурный оптимум, минимум и максимум.
Температурный оптимум- температура, при которой данный вид микробов наиболее хорошо развивается, т.е. температура, соответствующая физиологическим требованиям соответствующего микроорганизма. При температурном минимуме или максимуме развитие микробов еще возможно, но уже ограничено. При температуре выше максимума микробы обычно погибают. При температуре ниже минимума они переходят в состояние анабиоза, а при повышении температуры могут возвращаться к активной жизни.
По отношению к температурному фактору микроорганизмы делят на три группы – психрофилы (холодолюбивые), мезофилы ( развивающиеся при средних температурах) и термофилы (теплолюбивые) Такое деление производят на основе оптимальной температуры развития. Примерные границы температур для различных групп представлены в таблице 1.
Таблица 1 Температуры для различных групп микроорганизмов, °С
| Микроорганизмы | Минимальная | Оптимальная | Максимальная |
| Психрофилы | - 8 - 10 | + 10 + 15 | + 15 + 20 |
| Мезофилы | + 5 + 10 | + 30 + 37 | + 40 + 45 |
| Термофилы | + 15 + 20 | + 40 + 55 | + 60 + 70 |
Вышеуказанные температурные границы приведены для размножения микроорганизмов. Для других процессов жизнедеятельности (спорообразование, образование токсинов, пигментов и др.) значения температур для тех же групп микроорганизмов могут быть иными.
Психрофилами - называют микроорганизмы, область температур роста которых лежит в пределах от 0 (или ниже) до 20 °С, хотя оптимум составляет 15°С. Психрофильные микроорганизмы являются обитателями холодных источников, глубоких озер и океанов, хорошо развиваются на продуктах при холодильном хранении. Наиболее сильной устойчивостью к низким температурам обладают плесневые грибы и гнилостные бактерии (-3-9°С).
Мезофилы живут при средних температурах. Самая распространенная группа микроорганизмов (бактерии, плесневые грибы, дрожжи). Мезофилами являются все патогенные и условно-патогенные микроорганизмы и большинство сапрофитных.
Термофилы развиваются при высоких температурах. Они в большом количестве встречаются в почве, сточных водах и в навозе, в гейзерах, песках пустынь. Они участвуют в ряде биологических процессов: при самосогревании влажного сена и хлопка, силосовании кормов, вызывают порчу пастеризованных и стерилизованных продуктов. Знание отношения разных видов микробов к воздействию температур позволяет культивировать их в лабораториях на искусственных питательных средах. При этом учитывают значения оптимальных для каждого вида микробных клеток температурных режимов (в термостатах).
Отношение микроорганизмов к различным температурам стали использовать для сохранения различных пищевых продуктов. При этом используют как низкие, так и повышенные температуры. На основе этого применяют несколько технологических приемов обработки и хранения продуктов. Низкие температуры - хранение в охлажденном состоянии и замороженном. При хранении в охлажденном состоянии используют температуру 0 -+4°С, что позволяет продлить срок хранения, но если субстрат (продукт) достать из холодильника и оставить при комнатной температуре – он быстро испортится за счет развития тех микроорганизмов, что находились в нем до охлаждения. При хранении продуктов в замороженном состоянии используют температуру -12 - 30°С. Несмотря на то, что при таких температурах микроорганизмы не размножаются и активная деятельность их приостанавливается, многие из них неопределенно долгое время остаются жизнеспособными, переходя в анабиотическое состояние. При хранении продуктов в охлажденном и замороженном состоянии большое значение имеет относительная влажность воздуха, скорость охлаждения и замораживания, исходная степень обсеменения психрофильными микроорганизмами. Замораживание не оказывает стерилизующего действия и могут выжить многие виды сапрофитов и болезнетворные формы микроорганизмов. Поэтому размороженные продукты могут быстро подвергаться порче. Размораживать замороженные продукты следует непосредственно перед употреблением.
В пищевой промышленности применяют два способа воздействия высоких температур: пастеризация и стерилизация.
Пастеризация – это нагревание продукта чаще при температуре 63-80 °С в течение 20-40 мин. Иногда пастеризацию проводят кратковременно в течение нескольких секунд при температуре 90-100 °С. При пастеризации погибают не все микроорганизмы. Некоторые термоустойчивые бактерии и споры грибов остаются жизнеспособными. Поэтому пастеризованные продукты следует немедленно охлаждать дотемпературы не выше 10 °С и хранить на холоде ( на льду и в холодильнике), чтобы задержать прорастание спор и развитие сохранившихся клеток. Пастеризуют молоко и молочные продукты, пиво, соки, рыбную икру, пресервы и некоторые другие продукты.
Стерилизация - это температура 112-120 °С в течение 20-60 мин. в специальных приборах - автоклавах (перегретым паром под давлением) или при 160-180°С в течение 1-2 часа в сушильных шкафах (сухим жаром).
Влажность. Микроорганизмы могут развиваться только в субстратах, имеющих свободную воду и в количестве не менее определенного уровня. С понижением влажности субстрата интенсивность размножения микробов замедляется, а при удалении из субстратов ниже необходимого уровня вообще прекращается. Потребность во влаге у различных микроорганизмов колеблется в широких пределах. По величине минимальной потребности во влаге для роста различают следующие группы: гидрофиты (влаголюбивые), мезофиты (средневлаголюбивые), ксерофиты (сухолюбивые). Гидрофитами являются большинство бактерий, а мицелиальные грибы и дрожжи мезофиты, но имеются среди них и гидрофиты.
Для развития микроорганизмов имеет значение не абсолютная величина, а доступность содержащейся в субстрате воды, которую в настоящее время принято обозначать термином водная активность или аw. Водная активность показывает отношение давления водяных паров раствора (субстрата ) Р и чистого растворителя (воды) Ро при одной и той же температуре: а w = Р/Ро.
Водная активность выражается величинами от 0 до 1 и характеризует относительную влажность субстрата. Рост микроорганизмов наблюдается при значениях аw от 0,99 до 0,65-0,61. Оптимальное значение для многих от 0,99-0,98, примерно в этих пределах находится водная активность скоропортящихся пищевых продуктов (мяса, рыба, плоды, овощи).
Бактерии развиваются при водной активности субстрата 0,94-0,90. Дрожжи - 0,88-0,85, мицелиальные грибы - 0,8. Но некоторые виды бактерий, дрожжей, мицелиальных грибов могут расти при водной активности -0, 75-0,62 (хотя и медленно).
Таким образом, продукты, у которых водная активность менее 0,7 , могут длительно сохраняться без микробной порчи. Перспективно, с точки зрения увеличения срока хранения скоропортящихся продуктов, искусственное снижение в них водной активности. Возможно снижение аw при добавлении специфических веществ, способных связывать воду.
Давно применяется хранение различных пищевых продуктов в сухом виде. В высушенном состоянии многие микробы сохраняют жизнеспособность в течение длительного времени. Устойчивы к высушиванию многие дрожжи и особенно споры бактерий и мицелиальных грибов (сохраняют способность к прорастанию десятки лет). Патогенные микробы (стафилококки, микрококки, брюшно-тифозные бактерии) могут сохраняться в сухом субстрате неделями и месяцами.
Для сохранения сухих продуктов без порчи большое значение имеют относительная влажность и температура в складских помещениях. Продукты обладают гигроскопичностью (могут отдавать влагу или поглощать ее). Между влажностью воздуха и влажностью продукта устанавливается определенное подвижное равновесие. При одной и той же относительной влажности воздуха различные продукты могут иметь разную равновесную влажность. Большинство бактерий способно развиваться в субстратах при равновесной относительной влажности воздуха в пределах не ниже 95-90%. Для дрожжей минимум в субстрате соответствует 90-85% относительной влажности воздуха, для большинства мицелиальных грибов - 80%, а для некоторых ксерофитных видов пределом является относительная влажность воздуха- 75-65%
Таким образом, возможность развития микроорганизмов в продуктах в связи с их влажностью можно учитывать как по величине водной активности продукта, так и по относительной влажности воздуха. Значение аw, умноженное на 100, соответствует относительной влажности воздуха, выраженной в процентах, когда система продукт - воздух находится в равновесии.
Относительная влажность воздуха изменяется от температуры: с понижением температуры воздуха уменьшается его влагоудерживающая способность и наоборот. Поэтому при снижении температуры в процессе хранения это приводит к увлажнению поверхности продукта, что способствует развитию находящихся на нем микробов. При хранении и перевозке высушенных продуктов необходимо принимать меры для предупреждения изменения их влажности.
При сублимационной сушке (высушивание под высоким вакуумом в замороженном состоянии) качество и пищевая ценность продуктов сохраняются значительно лучше, но микроорганизмы хорошо переносят такое высушивание и сохраняются жизнеспособными. Поэтому к таким продуктам следует предъявлять строгие санитарно-гигиенические требования.
Концентрация растворенных веществ и осмотическое давление. Внутриклеточное осмотическое давление обусловлено концентрацией растворенных веществ в цитоплазме клетки. У разных микроорганизмов оно колеблется в широких пределах и этим объясняется тот факт, что различные микроорганизмы могут обитать в пресной воде и соленых водах морей. Высокие концентрации осмотически активных веществ способствуют плазмолизу микробных клеток. В качестве осмотически деятельных веществ, применяемых для консервирования пищевых продуктов, используют поваренную соль и сахар.
Большинство бактерий мало чувствительно к концентрации NаСL в пределах 0,5-2%, но 3% - ное ее содержание в среде неблагоприятно для многих микроорганизмов. Размножение многих гнилостных бактерий подавляется при концентрации поваренной соли 3-4%, а при 7-10% оно прекращается. Палочковидные гнилостные бактерии менее стойки, чем кокки. Развитие некоторых возбудителей пищевых отравлений (ботулинуса, сальмонелл) приостанавливается при 6-10% соли, но некоторые из них могут долго сохранять жизнеспособность даже при 20%. Микроорганизмы, нормально развивающиеся при высоких концентрациях поваренной соли (20% и выше) называют галофилами (солелюбивыми).
Концентрация соли, влияющая на развитие микроорганизмов, зависит от других условий среды (рН, температура). Развитие дрожжей в соленых продуктах подавляется в кислой среде при содержании соли 14%, а в нейтральной только при 20%. При понижении температуры подавляющее действие соли усиливается. При температуре 0 °С и 8% соли угнетается рост мицелиальных грибов, а при 20°С необходимо 12% соли для такого же эффекта. Имеются сведения об усилении действия поваренной соли в присутствии нитратов и нитритов.
Подавляющее воздействие соли на рост микроорганизмов объясняется не только повышением осмотического давления. Поваренная соль оказывает токсическое действие на микроорганизмы: подавляются процессы дыхания, нарушаются функции клеточных мембран и др.
Поскольку многие микроорганизмы в плазмолизированном состоянии длительное время не погибают, приостанавливается лишь их активная деятельность, к перерабатываемому сырью необходимо предъявлять строгие санитарно-гигиенические требования. Порча соленых товаров (мясо, рыба и др.) часто возможна под влиянием галофильных и солеустойчивых микроорганизмов. Например, покраснение крепко соленой рыбы - «фуксин», вызывается галофильной бактерией, обладающей красным пигментом. Для задержки развития микроорганизмов соленые товары необходимо хранить при низких температурах.
Возможны различные виды порчи (плесневение, забраживание меда, джема, варенья, фруктовых сиропов и других сахаросодержащих продуктов) под воздействием осмофильных плесеней и дрожжей. Порчу продуктов, прошедших тепловую обработку, вызывают осмофильные температуровыносливые дрожжи, но порча может явиться и результатом вторичного инфицирования продуктов микробами извне. Поэтому для предотвращения такого вида порчи необходимо разливать продукт в горячем виде в стерильную тару, герметично укупоривать ее и хранить при пониженной температуре.
Лучистая энергия. Различные формы лучистой энергии оказывают на микроорганизмы разнообразное физическое, химическое и биологическое действие. Биологическое действие излучения зависит от длины волны, чем она короче , тем в ней больше заключено энергии, тем сильнее воздействие на организм. В основе действия лежат физические и химические изменения, происходящие в клетках микроорганизмов и в окружающей среде. Изменения могут быть вызваны только поглощенными лучами. Следовательно, для эффективности действия излучения большое значение имеет проникающая способность лучей.
Солнечный свет обладает наибольшим потенциалом вредного воздействия на микроорганизмы. Способностью использовать энергию солнечного света обладают лишь пигментобразующие формы бактерий. Микроорганизмы, не имеющие пигмента, погибают под действием прямых солнечных лучей. Рассеянный солнечный свет подавляет их развитие постепенно. Однако, развитие многих мицелиальных грибов при постоянном отсутствии света протекает ненормально, хорошо развивается только мицелий, а спорообразование только тормозится. Под влиянием солнечных лучей происходят внутриклеточные химические реакции с образованием гидроксильных радикалов и других высокореактивных веществ, действующих губительно на микробную клетку.
Наиболее выраженное летальное действие оказывают световые волны, лежащие в ультрафиолетовой области спектра (длина волны менее 400 нм).
Ультрафиолетовые лучи (УФ - лучи) обладают или бактерицидным или мутагенным действием. Это вызывается изменениями в структуре ДНК. Из всех микроорганизмов наиболее чувствительны к УФ - лучам вегетативные формы бактерий, а споры бацилл в 4-5 раз более устойчивы. Очень чувствительны к УФ - лучам патогенные микроорганизмы .
Эффективность воздействия УФ - лучей зависит от дозы облучения, длительности и свойств облучаемого субстрата (рН, степень обсеменения микробами и температура). Очень малые дозы облучения действуют даже стимулирующе на отдельные функции микроорганизмов. Более высокие - могут вызвать изменение наследственных свойств. Это используется на практике для получения различных штаммов микроорганизмов с высокой способностью продуцировать антибиотики, ферменты и др. БАВ. Дальнейшее увеличение дозы приводит к гибели.
В настоящее время УФ - лучи довольно широко применяют для дезинфекции воздуха микробиологических боксов, холодильных камер и производственных помещений. Искусственным источником ультрафиолетового излучения служат аргонно-ртутные лампы низкого давления, называемые бактерицидными (БУФ-15, 30, 60). При обработке УФ - лучами в течение 6 часов уничтожается до 80% бактерий и мицелиальных грибов, находящихся в воздухе. Такие лучи могут быть использованы для предотвращения инфекции извне, при розливе, фасовке и упаковке пищевых продуктов, лечебных препаратов, а также для обеззараживания тары упаковочных материалов, оборудования, посуды (на предприятиях общественного питания).
Стерилизация пищевых продуктов с помощью УФ - лучей затруднена вследствие их невысокой проникающей способности. Действие их проявляется только на поверхности или в очень тонком слое. Предлагается использовать УФ - лучи для стерилизации плодовых соков и вин (в тонком слое). При таком «холодном» способе стерилизации вино поучается лучшего качества и сохранятся без порчи дольше, чем пастеризованное.
ля некоторых продуктов, таких как сливочное масло, молоко, стерилизация УФ - лучами неприемлема. В результате такой обработки ухудшаются вкусовые и пищевые свойства таких продуктов. В последнее время УФ - лучи используют для дезинфекции питьевой воды.
Космические и рентгеновские лучи представлены ионизирующими излучениями с длиной волны от 0,006 до10 нм. Они оказывают мутагенное или летальное действие. К действию таких лучей наиболее чувствительны ядерные структуры, хотя повреждаются и цитоплазматические структуры клеток.
Искусственное ионизирующее излучение (α- и b -частицы, g -лучи) возникает в результате работы атомных электростанций, испытаний ядерного оружия, применения радиоактивных изотопов в научных целях.
Эффект бактерицидного действия радиоактивных излучений обуславливается ионизацией внутриклеточных веществ. При прохождении ионизирующих излучений через клетку, некоторые атомы в результате поглощения энергии испускают электроны и превращаются в положительно заряженные ионы. Свободный электрон присоединяется к нейтральному атому, который превращается в отрицательно заряженный ион. Такое изменение электронной структуры атомов приводит к изменению химических связей и разрушению структур молекул.
Микроорганизмы более устойчивы к излучениям, чем высшие животные и растительные организмы. Дрожжи и плесени более устойчивы, чем бактерии. Споры бацилл и клостридий выносливее их вегетативных форм. Чувствительны к облучению кишечная палочка, протей, многие бактерии рода псевдомонас (распространенные возбудители порчи сырья и мясных и рыбных продуктов). Микрококки отличаются повышенной устойчивостью. Высока радиоустойчивость вирусов, у некоторых она превосходит устойчивость бактериальных спор. При одной и той же поглощенной дозе радиопоражаемость микроорганизмов одного и того же вида изменяется в зависимости от возраста клеток, состава среды, температуры, дозы и длительности облучения. Установлено, что микроорганизмы способны восстанавливать лучевые повреждения, что определяется видовыми особенностями микроорганизмов и их физиологическим состоянием. Искусственные ионизирующие излучения используют для стерилизации лечебных препаратов и некоторых пищевых продуктов (но при этом могут ухудшаться вкус и пищевые качества). Их используют для задержки прорастания картофеля и овощей, зерна и зернопродуктов, сухофруктов, а также для ускорения или замедления созревания плодов и в других целях. Наиболее приемлемы для этих целей гамма – лучи, обладающие наиболее высокой проникающей способностью и не вызывающие при облучении появления в продукте «наведенной» радиации.
Радиоволны. Короткие электромагнитные волны длиной от 10 до 50 м, ультракороткие длиной от10м до мм обладают стерилизующим эффектом. При прохождении коротких и ультракоротких радиоволн через среду возникают переменные токи высокой частоты (ВЧ) и сверхвысокой частоты (СВЧ). В электромагнитном поле электрическая энергия преобразуется в тепловую.
Характер нагревания в СВЧ поле отличается от характера нагрева от обычных нагреваний и обладает рядом преимуществ. Объект нагревается быстро и равномерно по всей массе. Например, воду в стакане можно довести до кипения в течение двух – трех секунд. Рыба (1кг) варится до готовности в течение двух мин., мясо (1кг)-2,5 мин., курица 6-8 мин. Нагрев может происходить избирательно, отдельные части облучаемого объекта нагреваются в разной степени и зависят от их электрофизических свойств.
Благодаря специфическим особенностям перспективно применение этого способа нагревания для пастеризации и стерилизации пищевых продуктов (например, в плодово-ягодных консервах). По сравнению с обычной паровой стерилизацией в автоклавах время нагревания СВЧ- энергией до одной и той же температуры сокращается во много раз. Поэтому полнее сохраняются вкусовые и питательные свойства. Эффект воздействия на его микрофлору практически одинаков.
Сверхвысокочастотную электромагнитную обработку пищевых продуктов все шире применяют в общественном питании (для варки, сушки, выпечки, при разогревании др.).
Давление и механическое сотрясение . Микроорганизмы не испытывают значительных изменений под влиянием даже очень больших давлений, но есть группы микроорганизмов, которые развиваются только при избыточных давлениях. Их называют барофильными (в глубинах морей и океанов). К механическим сотрясениям они чувствительны, если они сильные и длительные. Так, самоочищение бурных рек происходит в результате гибели микроорганизмов под воздействием сильных толчков воды.
Ультразвук. Ультразвуком называют механические колебания с частотами более 20000 колебаний в секунду (20 кГЦ). Колебания такой частоты находятся за пределами слышимости человека. Ультразвуковые волны могут распространяться в твердых, жидких и газовых средах. Обладают большой механической энергией и вызывают ряд физических, химических и биологических явлений. Механизм бактерицидного действия ультразвука объясняется двумя теориями: кавитационно - механической и кавитационно - электрохимической. По первой теории считают, что ультразвуковые волны, распространяясь в упругой среде, вызывают в ней попеременные сжатия и разряжения. В клетке создаются огромные давления, достигающие десятков и сотней мПа, что вызывает механическое разрушение цитоплазматических структур и гибель клетки (кавитация).
Кавитационная электрохимическая теория объясняет ионизацию паров жидкостей и присутствующих в ней газов при образовании кавитационного пузырька. При разрыве пузырька происходит электрический разряд, сопровождающийся резким повышением температуры и образованием в кавитационной полости электрического поля высокого напряжения. При этом пары жидкости и высокомолекулярные соединения в кавитационной полости расщепляются на водород и гидроксильную группу с образованием активного кислорода, перекиси водорода, азотистой и азотной кислот, в результате чего происходят инактивация ферментов и коагуляция белков. Все это обуславливает гибель микробной клетки.
Эффективность действия УЗ при одной и той же интенсивности и частоте колебаний зависит от продолжительности воздействия, химического состава облучаемой среды, ее вязкости , температуры, рН и исходной степени обсемененности микроорганизмами. Чем больше микроорганизмов, тем продолжительнее должно быть воздействие для достижения стерилизующего эффекта.
Устойчивость микроорганизмов к действию ультразвука зависит от их биологических свойств. Вегетативные клетки более чувствительны, чем споры, кокковые формы погибают медленнее, чем палочковидные. Более крупные клетки микроорганизмов отмирают быстрее, чем мелкие. Ультразвук применяют для стерилизации пищевых продуктов (молоко, фруктовые соки, вина), изготовления вакцин, мойки и стерилизации стеклянной тары, а также при извлечении внутриклеточных ферментов, токсинов, витаминов, нуклеиновых кислот и других компонентов клетки. Ведутся исследования по применению УЗ энергии для стерилизации питьевой воды.
3 Химические факторы. К химическим факторам, влияющим на жизнедеятельность микробов, относят: химический состав питательной среды, реакцию среды, окислительно-восстановительный потенциал среды и действие ядовитых (антисептических) веществ.
Состав питательной среды является основным показателем микроорганизмов. Он определяет ее питательную ценность, реакцию (рН) и окислительно-восстановительный потенциал (Еh).
Реакция питательной среды (концентрация водородных ионов рН) играет роль фактора, определяющего границы существования живой материи. Кислотность среды воздействует на ионное состояние, а поэтому на доступность для организма многих химических веществ.
Ионы водорода влияют на электрический заряд коллоидов клеточной стенки. При сдвиге рН в кислую или щелочную сторону изменяется знак заряда поверхности клетки, что приводит к изменению ее проницаемости для различных молекул и ионов питательного субстрата и нарушению нормального процесса обмена веществ. Изменение рН также влияет на степень дисперсности коллоидов цитоплазмы, активность ферментов, интенсивность и направление биохимических реакций. Так, например, дрожжи в кислой среде (рН 4,5-5,0) образуют в основном этиловый спирт (спиртовое брожение), а в щелочной среде (рН 8,2) – глицерин (глицериновое брожение).
В зависимости от отношения микрооргаизмов к кислотности среды их подразделяют на ацидофилы (кислотолюбивые), нейтрофилы (нейтральная зона) и алкалофилы (щелочелюбивые). Микроорганизмы, обладающие способностью выживать при значениях рН за пределами 4-9, рассматриваются как кислото - и щелочетолерантные.
Кислотолюбивые микроорганизмы, растущие при очень низком значении рН, встречаются редко. К ацидофильным относятся уксуснокислые, молочнокислые, некоторые дрожжи и плесени. Уксуснокислые бактерии растут в пределах рН от 3 до 5, молочнокислые развиваются при рН от 3 до 8. Оптимум рН роста дрожжей находится в области 4,5 - 6. Однако некоторые из них способны развиваться в более кислой среде – рН 2, другие - в щелочной - 8,5.
К самым устойчивым к кислой среде относятся плесневые грибы, многие из них характеризуются ацидотолерантностью и способностью роста в широких пределах рН (от 2 до11).
Оптимальная рН для нейтральнофильных микроорганизмов находится в пределах 7,0. Типичными представителями нейтрофилов являются бактерии группы кишечных палочек (БГКП), стрептококки, бациллы, сальмонеллы и большинство других патогенных микроорганизмов.
К алкалофилам относят некоторые виды бактерий и мицелиальных грибов. Клубеньковые бактерии рода Ризобиум (Rhizobium) активно развиваются при рН 10-12. Бациллюс цереус (Bacillus cereus) и Бациллус циркулянс (Bacillus circulans) способны развиваться при рН 10-11. Энтерококки также толерантны к щелочной среде.
Многие микроорганизмы, развиваясь в питательной среде, выделяют продукты обмена, изменяющие реакцию субстрата, это является одним из факторов, обусловливающих антогонизм между различными группами микробов. Так, молочнокислые бактерии в процессе жизнедеятельности образуют молочную кислоту, которая подавляет развитие большинства гнилостных бактерий. Это используется при хранении кисломолочных продуктов, сыров, при консервировании силоса, квашении капусты и других продуктов. Зная отношение различных микроорганизмов к реакции среды и регулируя ее рН, можно подавлять или стимулировать их развитие.
Окислительно-восстановительный потенциал служит количественной мерой способности некоторых соединений или элементов отдавать электроны. Этот отсчитывается относительно потенциала молекулярного водорода. Окислительно-восстановительные условия питательной среды выражаются величиной окислительно-восстановительного потенциала, который принято обозначать Еh (rH2). Окислительно-восстановительный (ОВ) потенциал среды представляет собой отрицательный логарифм числа, выражающего давление (в МПа) молекулярного водорода при давлении Н2 0,1 МПа. ОВ потенциал среды равен 0. Величина Еh минимальна при насыщении среды водородом и максимальна при насыщении ее кислородом. Она измеряется от 0 до 41 единиц. При равновесии окислительно-восстановительных процессов в среде rH2 равен 28. Присутствие в среде окисляющих веществ (метиленового синего, резазурина, кислот, перманганата калия и др.) повышает значение потенциала. Значение же соединений, обладающих восстановительными свойствами (цистеин, тимоловая кислота), снижает потенциал. ОВ потенциал также резко уменьшается при отмирании микроорганизмов, лизисе их фагом и действии на культуры лизоцимом. Изменяя ОВ потенциал среды, можно повлиять на интенсивность размножения различных групп микроорганизмов и направленность вызываемых ими биохимических процессов.
По отношению к окислительно-восстановительным условиям среды микроорганизмы разделяют на четыре основные группы: облигатные аэробы, облигатные и факультативные - анаэробы и микроаэрофилы. Облигатные анаэробы развиваются при низком значении Еh (0-14), факультативные анаэробы при Еh (0-30), аэробные микроорганизмы Еh (11-35), микроаэрофилы- Еh (10-20).
Влияние антисептических веществ на микробную клетку может проявляться различным действием. Одни подавляют жизнедеятельность или задерживают развитие чувствительных к ним микробов. Такое действие называют бактериостатическим (в отношении бактерий) или фунгистатическим (в отношении мицелиальных грибов). Другие вещества вызывают гибель микроорганизмов, оказывая на них соответственно бактерицидное или фунгицидное действие. В очень малых дозах многие химические яды оказывают даже благоприятное действие, стимулируя размножение или биохимическую деятельность микробов. В каждом конкретном случае доминирующий эффект зависит от химической природы этого антимикробного агента.
Эффективность действия химических веществ на микроорганизмы зависят от природы вещества, концентрации, биологических особенностей микроорганизмов, продолжительности воздействия, температуры, состава и рН среды. Чувствительность микроорганизмов к одному и тому же антисептику неодинакова.
Из неорганических соединений сильными ядами для микробов являются: соли тяжелых металлов (свинца, меди, цинка, серебра, золота, ртути),
различные окислители (хлор, хлорная известь, хлорамин, йод, бром, перманганат калия, пероксид водорода, озон, диоксид углерода, аммиак и др.),
минеральные кислоты (борная, серная, хлористоводородная, азотная и др.),
щелочи (гидроксид натрия, гидроксид калия и др.).
Среди органических соединений губительное воздействие оказывают органические кислоты (молочная, салициловая, масляная, уксусная, бензойная и др.), используемые в качестве консервантов в пищевой и парфюмерно-косметической промышленностях. Консервант должен обладать определенными липофильными свойствами для того, чтобы проникать через гидрофобную клеточную оболочку или разрушать ее. В то же время, для антимикробного действия консерванту требуется хорошая растворимость в воде, так как развитие микроорганизмов происходит исключительно в водной фазе и поэтому консервант должен находиться именно в ней.
Эффективность конкретного консерванта неодинакова в отношении плесневых грибов, дрожжей и бактерий, то есть он не может быть эффективен против всего спектра возможных микроорганизмов, вызывающих порчу пищевых продуктов.
Кроме того, к дезинфицирующим веществам этой группы относятся также диэтиловый эфир, спирты жирного и ароматического ряда (этиловый, бутиловый, амиловый, пропиловый и др.), эфирные масла, смолы, дубильные вещества, органические красители, формалин, фенол, крезол и их производные..
Ионы серебра и золота обладают олигодинамическим действием. В очень малых количествах, не поддающихся химическому обнаружению, они губительно действуют на микробные клетки. На этом основан метод дезинфекции воды с помощью серебряных фильтров. Посуда из серебра при контакте с водой сообщает ей бактерицидные свойства, этим объясняется длительное хранение «святой воды».
Химические вещества, бактерицидно действующие на микроорганизмы в небольших концентрациях, называют антисептическими или дезинфицирующими. Механизм бактерицидного действия антисептических веществ заключается в том, что в результате взаимодействия химического яда с веществами цитоплазмы в ней происходят необратимые изменения, вызывающие нарушения процессов жизнедеятельности и приводящие к гибели клетки.
Соли тяжелых металлов вызывают коагуляцию белков клетки. Олигодинамическое действие серебра и др. тяжелых металлов заключается в том, что положительно заряженные ионы металлов адсорбируются на отрицательно заряженной поверхности микробов, изменяется проницаемость их цитоплазматической мембраны и при этом нарушаются процессы питания и размножения микроорганизмов.
Окислители действуют на сульфгидрильные группы активных белков и влияют на другие группы (феноловые, тиоловые, индольные и аминные).
Неорганические кислоты и щелочи гидролизуют белки клетки. Диоксид углерода, сероводород, цианистые соединения инактивируют ферменты клетки.
Органические спирты, диэтиловый эфир, ацетон разрушают полипептидную оболочку клетки. Формалин (40%-й раствор формальдегида) присоединяется к аминогруппам белков и вызывает их денатурацию.
Многие антисептические вещества используются в медицине, сельском хозяйстве, в промышленности и в быту, как дезинфицирующие средства для борьбы с болезнетворными микробами. Широко применяют хлор и его соединения для дезинфекции питьевой воды, тары, оборудования, инвентаря.
Антисептические вещества используют для защиты от микробных поражений текстильных материалов, древесины, бумаги и изделий из нее, других материалов и объектов.
Применение антисептиков для консервирования продуктов ограничено и строго нормируется санитарным законодательством.
В нашей стране разрешено использовать немногие химические консерванты в малых дозах (от сотых до одной двух десятых процента) и только для некоторых пищевых продуктов.
Для консервирования полуфабрикатов из плодово-ягодного сырья, рыбных консервов, кетовой икры используют бензойную кислоту и ее натриевую соль. В качестве консерванта для многих пищевых продуктов все чаще применяют сорбиновую кислоту и ее соли. Эта кислота менее токсична, чем бензойная и сернистая, и более активно воздействует на микроорганизмы. При концентрации 0,03-0,1% эта кислота на длительное время задерживает рост грибов, дрожжей и некоторых бактерий и при этом безвредна для людей, не придает продукту посторонних вкуса и запаха. Особенно эффективно действие сорбиновой кислоты в кислой среде рН 3-4,5. Этот консервант вводят непосредственно в продукт или обрабатывают им поверхность продукта, оберточные материалы.
Для борьбы с картофельной болезнью хлеба, для предотвращения его плесневения рекомендуется введение в тесто солей пропионовой кислоты. Этот консервант можно применять и для некоторых рыбных продуктов.
4 Биологические факторы. Под биологическими факторами понимают влияние на жизнедеятельность микроорганизмов других видов и групп микробов, а также животных и растений, составляющих в природных условиях специфический биоценоз. В процессе эволюции возникли и сформировались различные типы взаимоотношений между микроорганизмами. В связи с этим различают несколько типов взаимоотношений (симбиоза) между организмами: мутуализм, синергизм, комменсализм, паразитизм, метабиоз и антагонизм (антибиоз).
Мутуализм (взаимовыгодный симбиоз) представляет собой сожительство, благоприятное для обоих симбионтов, совместно они развиваются даже лучше, чем каждый в отдельности. Примером может служить совместное развитие молочнокислых бактерий и дрожжей (в кефирных грибках).Молочнокислые бактерии в кефирных грибках продуцируют молочную кислоту и создают среду, благоприятную для роста дрожжей, а дрожжи, выделяя витамины группы В, стимулируют развитие молочнокислых бактерий. Симбиотические взаимоотношения этих микроорганизмов используют в процессе изготовления некоторых молочнокислых продуктов (кефира, кумыса).
Синергизм - содружественное действие двух или нескольких видов, когда при совместном развитии усиливаются отдельные физиологические функции. Например, повышается синтез определенных веществ (образование ароматических веществ лактококками при совместном выращивании с молочнокислыми стрептококками.
Комменсализм – тип взаимоотношений между двумя организмами, при котором один живет за счет другого, не принося заметной пользы и не причиняя вреда. Такие взаимоотношения наблюдаются между молочнокислыми бактериями, а также кишечными палочками и организмом человека или животного. При развитии в толстом отделе кишечника бактерии получают от макроорганизма необходимые питательные вещества, не причиняя ему вреда и даже принося известную пользу тем, что подавляют развитие гнилостных и некоторых патогенных микроорганизмов.
Паразитизм – вид взаимоотношений, когда один из них (паразит) живет за счет другого (хозяина), причиняя ему вред. Паразитами являются все патогенные микроорганизмы по отношению к человеку, животному и растениям. Абсолютными паразитами являются риккетсии и вирусы, развивающиеся внутри клеток макро- и микроорганизмов.
Метабиоз - такой вид взаимоотношений, когда продукты жизнедеятельности одного микроорганизма являются продуктами питания других. Так, дрожжи, сбраживая сахар в этиловый спирт, создают условия для развития уксуснокислых бактерий, а образуемая последними уксусная кислота используется плесенями, которые ее окисляют до С2О и Н2О.
Антагонизм (антибиоз) – тип взаимоотношений между микроорганизмами, при котором одни микроорганизмы подавляют развитие других. Причин антагонизма может быть несколько: истощение питательного субстрата вследствие более быстрого развития одного из микроорганизмов; изменение рН среды (при развитии ацидофилов, алкалофилов); выделение в среду микробами - антагонистами антибиотиков. Антибиотики - вещества биологического (микробного, растительного и животного) происхождения, подавляющие развитие и биохимическую активность чувствительных к ним микробов. По происхождению антибиотики подразделяют на группы: антибиотические вещества, продуцируемые актиномицетами, плесневыми грибами, бактериями, организмом животного или человека; антибиотики растительного, синтетического и полусинтетического происхождения Известно более 5000 антибиотиков, обладающих различным спектром действия.
Антибиотики актиномицетного происхождения - стрептомицин, тетрациклины, неомицин, нистатин обладают широким антибактериальным спектром действия. Они активны в отношении грамположительных бактерий, возбудителей туберкулеза, брюшного тифа, туляремии, бруцеллеза, сальмонеллезов и др.
Наиболее активными продуцентами антибиотиков являются мицелиальные грибы. Плесень рода Penicillium продуцирует широко используемый пенициллин. Он обладает бактерицидным действием главным образом на грамположительные стафилококки и стрептококки. Плесени рода Aspergillus выделяют антибиотики- фумингацин и аспергиллин. Mucor продуцирует клавицин.
К антибиотикам, продуцируемым бактериями, относят грамицидин – С (Вac. Brevis), пиоцианин (Ps. aeruginoza), субтилин (Bac. subtilis), полимиксин (Bac. polymixa). Молочнокислые бактерии Lbm. Plantarum выделяют антибиотик лактолин, Lac. Lactis –низин, Lac. Cremoris- диплококцин и др. Эффективность бактериальных антибиотиков ниже, чем антибиотиков грибного и актиномицетного происхождения, однако они способны подавлять развитие возбудителя туберкулеза, маслянокислых бактерий, кишечных палочек, стафилококков и других видов молочнокислых бактерий.
К антибиотическим веществам животного происхождения относят лизоцим, эритрин и экмолин. Лизоцим содержится в яичном белке, слезах, слюне, молозиве, молоке. Он убивает и растворяет (лизирует) многие виды бактерий. Эритрин получен из красных кровяных шариков (эритроцитов) крови животных, проявляет бактериостатическую активность. Экмолин получают из тканей рыб. Он активен в отношении стафилококков и стрептококков.
Антимикробные вещества высших растений называют фитонцидами. Наиболее сильной бактерицидностью обладают фитонциды лука, чеснока, хрена, горчицы, алое, крапивы, можжевельника, почек березы, листьев черемухи и др. Антимикробное действие фитонцидов обусловлено продуктами жизнедеятельности растительных организмов: эфирных масел, глюкозидов, органических кислот, дубильных веществ, смол и др.
Полусинтетические антибиотики получают химическим путем. Они имеют широкий спектр действия, активны в отношении не только грамположительных, но и грамотрицательных микроорганизмов (исключение составляет синегнойная палочка). Синтезированы полусинтетические пенициллины (оксациллин, ампициллин, карбенициллин), цефалоспорины (цефалоредин), тетрациклины (метацилиногидрохлорид) и др.
Химическая природа антибиотиков различна. Они отличаются химической структурой и биологическими свойствами. Антибиотические вещества из бактерий являются полипептидами, а выделенные из актиномицетов и грибов относятся к сложным циклическим соединениям.
Лекция № 10 Важнейшие биохимические процессы превращений, вызываемые микроорганизмами: брожения - спиртовое, молочнокислое, масляно-кислое и их модификации; окислительные брожения - уксусно-кислое, лимонно-кислое; превращение белков - гниение.
Луи Пастером установлено, что изменения, происходящие при брожении, являются результатом жизнедеятельности микроорганизмов. В зависимости от преобладающих конечных продуктов различают типы брожений: спиртовое, молочно-кислое, масляно-кислое, их модификации и др. виды брожения.
Спиртовое брожение. Спиртовым брожением называют процесс превращения сахаров под действием ферментативной активности некоторых видов бактерий, мицелиальных и дрожжевых грибов с накоплением в качестве основного продукта этилового спирта и углекислого газа:
С6Н12О6®2С2Н5ОН + 2СО2+ Q
Но основными возбудителями этого вида брожения являются дрожжи (сахаромицеты). В анаэробных условиях превращение сахара в спирт происходит не сразу, процесс идет через ряд промежуточных реакций и протекает как бы в две стадии: первая (окислительная) включает превращение глюкозы до пировиноградной кислоты с образованием двух молекул восстановленного НАД×Н2 (первичный акцептор водорода):
С6Н12О6®2СН3СОСООН+2 НАД×Н2.
Под действием фермента пируватдекарбоксилазы дрожжи катализируют реакцию декарбоксилирования пировиноградной кислоты с отщеплением СО2 и образованием уксусного альдегида:
2СН3СОСООН®2СН3СНО+ 2СО2,
а вторая стадия (восстановительная) - НАД×Н2 передает Н конечному акцептору (уксусный альдегид), который превращается в этиловый спирт:
2СН3СНО+ 2НАД×Н2 ®2С2Н5ОН+ НАД
С энергетической точки зрения брожение - процесс мало экономичный, так как при сбраживании грамм-молекулы глюкозы синтезируется всего два моля АТФ.
Наряду с главными продуктами брожения в небольшом количестве образуются побочные продукты: глицерин, уксусный альдегид, уксусная и янтарная кислоты, сивушные масла (смесь высших спиртов) и некоторые другие вещества.
Нормальное спиртовое брожение протекает в кислой среде при рН 4-5. В щелочной среде при подщелачивании среды до рН 8 или при введении в среду бисульфита натрия направление брожения изменяется в сторону увеличения выхода глицерина (глицериновое брожение):
2С6Н12О6®2СН2ОНСНОНСН2ОН +С2Н5ОН +СН3СООН + 2СО2+ Q
При интенсивной аэрации среды дрожжевые грибы меняют тип энергетического обмена и переходят с процесса брожения на процесс дыхания, что называют эффектом Пастера:
С6Н12О6 + 6О2 ®6СО2 +6Н2О + Q
Дыхание является более выгодным энергетическим процессом, так как при сбраживании грамм-молекулы глюкозы синтезируется 36 молекул АТФ.
Процесс спиртового брожения лежит в основе хлебопечения, бродильных производств (виноделие, пивоварение, производство этилового спирта), глицерина. Совместно с молочно-кислым брожением используется при получении некоторых молочно-кислых продуктов (кефира, кумыса), при квашении овощей. Эффект Пастера используется для получения большого количества биомассы дрожжей (хлебопекарные и кормовые).
Молочно-кислое брожение. Это процесс превращения сахара в молочную кислоту. По характеру брожения различают две группы молочно-кислых бактерий: гомоферментативные (типичные) и гетероферментативные (нетипичные).
Гомоферментативные бактерии образуют в основном (не менее 85-90%) молочную кислоту и очень мало побочных продуктов:
2С6Н12О6®2СН3СНОНСООН + Q
Гетероферментативные бактерии наряду с молочной кислотой образуют значительное количество других веществ:
2С6Н12О6®2СН3СНОНСООН +СООН (СН2)2 СООН+ С2Н5ОН + СН3СООН+ Q
Есть такие гетероферментативные молочно-кислые бактерии, которые кроме того продуцируют четырехуглеродные соединения ацетоин (СН3СНОНСООСН3) и диацетил (СН3СОСОСН3), обладающие своебразным приятным ароматом.
В зависимости от условий развития (рН, температуры, степени аэробности и др.) характер конечных продуктов брожения может меняться у одного и того же вида молочно-кислых бактерий. Процесс превращения глюкозы до пировиноградной кислоты у гомоферментативных молочно-кислых бактерий протекает по гликолитическому пути. Затем, ввиду отсутствия фермента пируватдекарбоксилазы у этих бактерий, пировиногрданая кислота не подвергается расщеплению и является конечным акцептором водорода под действием фермента лактатдегидрогеназы:
СН3СОСООН+ НАД×Н2. ®2СН3СНОНСООН + НАД
Превращение глюкозы гетероферментативными молочно-кислыми бактериями происходит по-иному, так как они отличаются набором ферментов. Из-за отсутствия у них фермента альдолазы изменяется начальный путь превращения глюкозы, и процесс протекает не по гликолитическому пути, а по пентозофосфатному. Все молочно-кислые бактерии имеют форму кокков и палочек, неподвижны, не образуют спор, грамположительны, являются факультативными анаэробами.
Молочно-кислые бактерии широко применятся в различных отраслях пищевой промышленности (особенно молочно-кислой). Большое значение эти бактерии имеют при квашении овощей, силосовании кормов, в хлебопечении при приготовлении ржаного хлеба, при производстве некоторых колбас и солено-вареных мясных изделий, при созревании слабо соленой рыбы, при получении молочной кислоты.
Спонтанно (самопроизвольно) возникающее молочно-кислое брожение в пищевых продуктах приводит к их порче: прокисание, помутнение, ослизнение.
Пропионово-кислое брожение. Это превращение сахара или молочной кислоты и ее солей под действием ферментативной активности пропионово-кислых бактерий в пропионовую и уксусную кислоты с выделением углекислого газа и воды:
3С6Н12О6 ® 4СН3СН2СООН+ 2СН3СООН+ 2СО2 + 2Н2О+Q;
3 СН3СНОНСООН ® 2СН3СН2СООН+ СН3СООН+ СО2 + Н2О+Q
Некоторые пропионово-кислые бактерии могут образовывать, кроме того, муравьиную, янтарную и изовалериановую кислоты. При пропионово-кислом брожении превращение глюкозы до пировиноградной кислоты протекает также по гликолитическому пути. В дальнейшем пировиноградная кислота, претерпевая ряд превращений, восстанавливается в пропионовую. Пропионово-кислые бактерии относятся к актиномицетам. Это неподвижные, бесспоровые, грамположительные палочки, слегка изогнутые, факультативные анаэробы.
Пропионово-кислое брожение является одним из важных процессов при созревании сычужных сыров.
Пропионовая кислота и ее соли служат ингибиторами мицелиальных грибов и могут быть использованы для предотвращения плесневения пищевых продуктов. Некоторые виды пропионово-кислых бактерий применяют для получения витамина В12.
Слизевое (декстрановое) брожение. Вызывают слизеобразующие бактерии, которые по своим свойствам близки к молочно-кислым и способны образовывать вокруг клеточной стенки капсулу, состоящую из слизистых веществ (полисахариды, полипептиды). Процесс идет в несколько этапов:
С12Н22О11+ Н2О + ®С6Н12О6 +С6Н12О6.
сахароза глюкоза фруктоза
С6Н12О6. ®(С6Н10О5)n+Н2О
глюкоза декстран
С6Н12О6®2СН3СНОНСООН +СООН (СН2)2 СООН+ С2Н5ОН + СН3СООН+ Q
фруктоза
Сбраживание глюкозы протекает по гетероферментативному молочно-кислому брожению.
Слизеобразующие бактерии вызывают ослизнение овощей, фруктов, хлебобулочных изделий (картофельная болезнь), сахарных сиропов, различных слабоалкогольных напитков и вина. Промышленного значения это брожение не имеет.
Масляно-кислое брожение. Этот процесс представляет собой превращение сахара масляно-кислыми бактериями в анаэробных условиях с образованием масляной кислоты, углекислого газа и водорода:
С6Н12О6. ®СН3СН2СН2СООН + 2 СО2 +2Н2 + Q
В качестве побочных продуктов могут накапливаться бутиловый спирт, ацетон, этиловый спирт, уксусная кислота.
При масляно-кислом брожении сахар претерпевает те же превращения, что и при спиртовом и гомоферментативном молочно-кислом брожениях, вплоть до образования пировиноградной кислоты. Пировиноградная кислота подвергается декарбоксилированию и при участии кофермента А расщепляется до ацетилКоА, СО2, Н2. Две молекулы образовавшегося двууглеродного соединения конденсируются при участии фермента карболигазы. Из синтезированного четырехуглеродного соединения в сложном цикле последоваельных превращений образуется масляная кислота:
СН3СОСООН®СН3СНО + СО2;
СН3СНО+ СН3СНО® СН3СН2СН2СООН
Масляно-кислые бактерии представляют собой подвижные, крупные спорообразующие грамположительные палочки, строгие анаэробы.
В природе это брожение имеет положительное значение как звено в цепи многообразных превращений органических веществ. В пищевом производстве часто приносит значительный ущерб, так как может вызвать порчу картофеля и овощей, вспучивание сыров, порчу консервов (бомбаж), прогоркание молока, масла, увлажненной муки и т.д. Могут вызывать порчу квашеных овощей при замедленном молочно-кислом брожении, образующаяся при этом масляная кислота придает продуктам острый прогорклый вкус и резкий неприятный запах.
Масляно-кислое брожение применяют для производства масляной кислоты, которая представляет собой бесцветную маслянистую жидкость с неприятным резким запахом, а эфиры масляной кислоты отличаются приятным ароматом, как, например: метиловый эфир имеет яблочный запах, этиловый - грушевый, амиловый – ананасовый. Их используют как ароматические вещества в кондитерской, парфюмерной промышленности и при изготовлении фруктовых напитков.
Брожение пектиновых веществ. В тканях растений, овощей и фруктов пектины входят в состав клеточных стенок и срединных пластинок между отдельными клетками в виде труднорастворимого протопектина. Под действием пектолитических ферментов микроорганизмов, развивающихся в растительном пищевом сырье, протопектин превращатся в растворимый пектин. Пектин разлагается с образованием галактуроновых кислот, углеводов (ксилозы, галактозы, арабинозы), метилового спирта и других веществ. Сахара сбраживаются бактериями по типу масляно-кислого брожения с образованием уксусной и масляной кислот, СО2 и Н2 . Все эти процессы приводят к мацерации (распаду) пораженных объектов и другим видам повреждений. Разложение пектиновых веществ может быть выражено следующей схемой:
С46Н68О40 + nН2О® СНО (СНОН)4СООН + С6Н12О6 + С5Н10О5
Пектин галактуроновая к-та галактоза ксилоза
+ С5Н10О5 + СН3ОН +СН3СООН
арабиноза
С6Н12О6® СН3СН2СН2СООН + СН3СООН +СО2 +Н2+ Q
С5Н10О5® СН3СН2СН2СООН + СН3СООН +СО2 +Н2О+ Q
Продукты распада пектиновой кислоты (галактоза, арабиноза и др.) подвергаются сбраживанию или окислению разнообразными микроорганизмами. При анаэробных условиях они сбраживаются масляно-кислыми бактериями, относящимися к роду клостридий. Пектиновое брожение наблюдается при мочке лубоволокнистых растений - льна, конопли, кенафа, джута, канатника и др. Для освобождения волокон клетчатки необходимо разложение пектина, что и происходит под действием пектиноразлагающих ферментов анаэробных бактерий.
Брожение клетчатки. Клетчатка (целлюлоза) является основным веществом клеточных стенок растений. Брожение клетчатки состоит из разложения ее в анаэробных условиях палочковидными споробразуюищими бактериями рода клостридий. Они гидролизуют клетчатку до глюкозы в результате активности ферментов целлюлазы (до целлобиозы) и целлобиазы (до глюкозы) (см.уравнение 1-ниже). Глюкозу они сбраживают с образованием уксусной, масляной, молочной, муравьиной кислот, этилового спирта, СО2 и Н2 (уравнение 2 ниже):
(С6Н10О5)n + nН2О ®С12Н22О11 ® С6Н12О6;
С6Н12О6® СН3СООН +СН3СН2СН2СООН+ СН3СНОНСООН + С2Н5ОН +НСООН +СО2+ Q
Среди этих бактерий есть мезофилы и термофилы, которые накапливают различные органические кислоты.
Окислительные брожения. Окислительное брожение- это процесс окисления органических веществ (спиртов, углеводов) под действием ферментативной активности микроорганизмов не до конечных продуктов, как при дыхании, а до органических кислот и спиртов, как при брожении.
Уксусно - кислое брожение. Это процесс окисления этилового спирта под действием ферментов уксусно - кислых бактерий до уксусной кислоты и воды. Окисление этилового спирта протекает в две стадии - сначала образуется уксусный альдегид, который далее окисляется до уксусной кислоты:
2С2Н5ОН +О2® 2 СН3СНО + 2Н2О;
2СН3СНО+О2® 2 СН3СООН
Уксусно-кислые бактерии - бесспоровые палочки, грамотрицательные, строгие аэробы. Среди них есть подвижные и неподвижные формы. Уксусно-кислые бактерии относят к двум родам бактерий: Глюконобактер (Gluconobacter) и Ацетобактер ( Acetobacter). Ацетобактер – перитрихи, способные окислять уксусную кислоту до СО2 и Н2О (переокисление).
Уксусно-кислым бактериям свойственна изменчивость формы клеток, в неблагоприятных условиях могут образовывать толстые длинные нити, иногда раздутые уродливые клетки.
Уксусно-кислое брожение лежит в основе получения уксуса для пищевых целей. Исходным сырьем может служить спиртовый раствор, разбавленное подкисленное плодово-ягодное или виноградное вино.
Лимонно-кислое брожение. Это процесс неполного окисления углеводов ферментативной активностью мицелиальных грибов с образованием органических кислот (лимонной, щавелевой и др.). Окисление глюкозы в лимонную кислоту можно представить следующим суммарным уравнением:
2С6Н12О6+3О2® 2С6Н8О7 +4 Н2О
Химизм протекает по гликолитическому пути до образования пировиноградной кислоты. Пировинограданая кислота через ацетил-coA включается в цикл Кребса (цикл ди- и трикарбоновых кислот), в котором происходит накопление яблочной, фумаровой, янтарной и лимонной кислот.
Наибольшее практическое значение имеет процесс получения лимонной кислоты, которую получают в настоящее время с помощью гриба Аспергиллус нигер (Aspergillus niger) в кислой среде. При изменении кислотности среды до нейтральной происходит накопление щавелевой кислоты.
Технические приемы биохимического получения лимонной кислоты в нашей стране были разработаны С.П. Костычевым В.С. Буткевичем. Используют поверхностный и глубинный (более перспективный) способы промышленного получения лимонной кислоты
Превращения белков - гниение . В метаболизме микроорганизмов азотсодержащие вещества подвергаются разнообразным превращениям. По случайно поверхностному сходству разные виды порчи пищевых продуктов нередко называют гниением. Однако гниение - это процесс глубокого разложения белковых веществ микроорганизмами.
Способность разлагать в той или иной степени белковые вещества свойственна многим микроорганизмам. Некоторые из них разлагают непосредственно белки, другие могут воздействовать только на более или менее простые продукты распада белковой молекулы, например на пептиды, аминокислоты и др.
Продукты разложения белков микробы используют для синтеза веществ своего организма, а также в качестве энергетического материала.
Химизм разложения белковых веществ. Гниение - сложный, многоступенчатый биохимический процесс, характер и конечный результат которого зависят от состава разлагаемых белков, условий процесса и видов вызывающих его микроорганизмов.
Белковые вещества не могут непосредственно поступать в клетки микроорганизмов, поэтому использовать белки могут только те микроорганизмы, которые обладают протеолитическими ферментами — экзопротеазами, выделяемыми клетками в окружающую среду.
Процесс распада белков начинается с их гидролиза. Первичными продуктами гидролиза являются пептоны и пептиды. Они расщепляются до аминокислот, которые являются конечными продуктами гидролиза.
Образующиеся в процессе распада белков различные аминокислоты используются микроорганизмами или подвергаются ими дальнейшим изменениям, например дезаминированию, в результате чего образуются аммиак и разнообразные органические соединения. Процесс дезаминирования может происходить различными путями. Различают дезаминирование гидролитическое, окислительное и восстановительное.
Гидролитическое дезаминирование сопровождается образованием оксикислот и аммиака. Если при этом происходит и декарбоксилирование аминокислоты, то образуются спирт, аммиак и углекислый газ:
 RCHNH
RCHNH  СOOН + Н О RСНOНСООН + NН
СOOН + Н О RСНOНСООН + NН  ;
;
RСНNН СООН + Н О RCH OH + NН + CO ;
При окислительном дезаминировании образуются кетокислоты и аммиак:
RСНNН СООН + ½ O RСОCOOН+NНз
При восстановительном деэаминировании образуются карбоновые кислоты и аммиак:
RСНNН СООН + 2Н RCH СООН + NН
Из приведенных уравнений видно, что среди продуктов разложения аминокислот в зависимости от строения их радикала (R) обнаруживаются различные органические кислоты и спирты. Так, при разложении аминокислот жирного ряда могут накапливаться муравьиная, уксусная, пропионовая, масляная и другие кислоты, пропиловый, бутиловый, амиловый и другие спирты. При разложении аминокислот ароматического ряда промежуточными продуктами являются характерные продукты гниения: фенол, крезол, скатол, индол - вещества, обладающие очень неприятным запахом. При распаде аминокислот, содержащих серу, получается сероводород или его производные - меркаптаны (например, метилмеркаптан СН SН). Меркаптаны обладают запахом тухлых яиц, который ощущается даже при ничтожно малых концентрациях.
Образующиеся при гидролизе белка диаминокислоты могут подвергаться декарбоксилированию без отщепления аммиака, в результате чего получаются диамины и углекислый газ. Например, лизин превращается в кадаверин:
 NH
NH 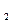 (CH )
(CH )  CHNH COOOH декарбоксилаза NH (CH )
CHNH COOOH декарбоксилаза NH (CH )  + CO
+ CO
Аналогично этому орнитин превращается в путресцин.
Кадаверин, путресцин и другие амины, образующиеся при гниении, часто объединяют под общим названием птомаины (трупные яды), некоторые из них обладают ядовитыми свойствами.
Дальнейшее превращение азотистых и безазотистых органических соединений, получающихся при распаде различных аминокислот, зависит от окружающих условий и состава микрофлоры. Аэробные микроорганизмы подвергают эти соединения окислению, так что они могут быть полностью минерализованы. В таком случае конечными продуктами гниения являются аммиак, углекислый газ, вода, сероводород, соли фосфорной кислоты. В анаэробных условиях не происходит полного окисления промежуточных продуктов распада аминокислот. В связи с этим, кроме аммиака и углекислого газа, накапливаются различные органические кислоты, спирты, амины и другие органические соединения, в числе которых могут быть вещества, обладающие ядовитыми свойствами, и вещества, придающие гниющему материалу отвратительный запах.
Возбудители гниения. Среди множества микроорганизмов, способных в той или иной мере разлагать белки, особое значение имеют микроорганизмы, которые вызывают глубокий распад белков - собственно гниение. Такие микроорганизмы принято называть гнилостными. Из них наибольшее значение имеют бактерии. Гнилостные бактерии могут быть спорообразующими и бесспоровыми, аэробными и анаэробными. Многие из них мезофилы, но есть холодоустойчивые и термостойкие. Большинство чувствительны к кислотности среды.
Наиболее распространенными и активными возбудителями гнилостных процессов являются следующие.
Сенная и картофельная палочки* - аэробные, подвижные, грамположительные, спорообразующие бактерии. Споры их отличаются высокой термоустойчивостью. Температурный оптимум развития этих бактерий 35 – 45 °С, максимум роста - при температуре около 50 - 55°С; при температуре ниже 5°С они не размножаются.
* В соответствии с Международным кодексом номенклатуры бактерий сенная и картофельная палочки ввиду большого сходства объединены в один вид - Васillus subtilis.
Помимо разложения белков, такие бактерии способны разлагать пектиновые вещества, полисахариды растительных тканей, сбраживать углеводы.
Сенная и картофельная палочки широко распространены в природе и являются возбудителями порчи многих пищевых продуктов. Они вырабатывают антибиотические вещества, подавляющие рост многих болезнетворных и сапрофитных бактерий.
Бактерии рода Рsеudоmоnas - аэробные подвижные палочки, с полярным жгутиком, не образующие спор, грамотрицательные (рисунок 2, а). Многие виды холодоустойчивы, минимальная температура их роста от - 2 до - 5°С, оптимум - около 20°С. Многие псевдомонасы помимо протеолитической обладают липолитической активностью; они способны сбраживать углеводы с образованием кислот, выделять слизь.
Развитие и биохимическая активность этих бактерий значительно тормозятся при рН ниже 5,5 и 5 - 6%-ной концентрации NаСl в среде. Псевдомонасы широко распространены в природе, являются антагонистами ряда бактерий и плесеней, так как образуют антибиотические вещества. Некоторые виды Рsеudоmоnas являются возбудителями болезней (бактериозов) культурных растений, плодов и овощей.
Протей (Ргоtеus vulgаris) - мелкие грамотрицательные бесспоровыс палочки с резко выраженными гнилостными свойствами. Белковые субстраты при развитии в них протея приобретают сильный гнилостный запах. В зависимости от условий жизни эти бактерии способны заметно менять свою форму и размеры (рисунок 2,6).
Протей - факультативный анаэроб; сбраживает углеводы с образованием кислот и газов. Он хорошо развивается как при температуре 25 °С, так и при 37°С, прекращая размножаться лишь при температуре около 5°С, однако может сохраняться и в замороженных продуктах.
Характерной особенностью протея является его очень высокая подвижность. Это свойство лежит в основе метода выявления протея на пищевых продуктах и отделения его от сопутствующих бактерий. Некоторые виды протея выделяют токсические для человека вещества.
Клостридиум путрификум (Сlоstridium putrificum) - анаэробная подвижная, спорообразующая палочка. Относительно крупные споры ее располагаются ближе к концу клетки, которая при этом приобретает сходство с барабанной палочкой. Споры довольно термоустойчивы. Углеводы эта бактерия не сбраживает. Белки разлагает с образованием большого количества газов (NН , Н 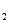 S). Оптимальная температура развития 37 – 43 °С, минимальная 5 °С.
S). Оптимальная температура развития 37 – 43 °С, минимальная 5 °С.
Клостридиум спорогенес (Сlоstridium sporogenes) - анаэробная подвижная спороносная палочка. Споры термоустойчивы, в клетке они расположены ближе к ее концу. Характерным является очень быстрое (в течение первых суток роста) образование спор. Эта бактерия сбраживает углеводы с образованием кислот и газа, обладает липолитической способностью. При разложении белков обильно выделяется сероводород. Оптимальная температура развития 35-40 °С, минимальная-около 5°С.
Оба вида клостридий известны как возбудители порчи баночных консервов (мясных, рыбных и др.).
Практическое значение процессов гниения. Гнилостные микроорганизмы наносят нередко большой ущерб народному хозяйству, вызывая порчу ценнейших и богатых белками продуктов питания, наприме, мяса и мясопродуктов, рыбы и рыбопродуктов, яиц, молока и др. Но эти микроорганизмы играют большую положительную роль в круговороте веществ в природе, минерализуя белковые вещества, попадающие в почву, воду.
Нитрификация. Процесс последовательного окисления аммиака до азотистой и азотной кислот называется нитрификацией, а возбудители его — нитрифицирующими бактериями. Сущность этого процесса была раскрыта и изучена С. Н. Виноградским.
Работами С. Н. Виноградского установлено, что процесс нитрификации происходит в две фазы, каждая из которых обусловлена деятельностью специализированных аэробных бактерий. Возбудители первой фазы — нитрозные бактерии - окисляют аммиак до солей азотистой кислоты (нитритов). Возбудители второй фазы - нитратные бактерии - окисляют соли азотистой кислоты в соли азотной кислоты (нитраты).
Нитрифицирующие бактерии относятся к хемоавтотрофам. Они живут в почве, природных водоемах, обогащая их нитратами - лучшим источником азотистого питания для растений.
Денитрификация. Процесс восстановления нитратов до молекулярного азота называется денитрификацией, а бактерии, осуществляющие его, — денитрифицирующими бактериями.
Денитрифицирующие бактерии - факультативные анаэробы. В аэробных условиях в процессе их дыхания конечным акцептором водорода является кислород. В анаэробных условиях в качестве акцептора водорода используются нитраты (некоторые используют нитриты), которые и восстанавливаются до молекулярного азота. Многие микроорганизмы восстанавливают нитраты только до нитритов.
Денитрифицирующие бактерии широко распространены в природе. Они живут в почве, природных водах. Деятельность денитрифицирующих бактерий в почве отрицательна, особенно при анаэробных условиях, так как азот нитратов, усваиваемый растениями, переходит в неиспользуемый ими свободный азот.
Фиксация молекулярного азота. Некоторые бактерии способны фиксировать свободный атмосферный азот, т. е. переводить его в связанное состояние. Они восстанавливают азот до аммиака; часть его используется самими микроорганизмами, а часть выделяется в окружающую среду.
Одни азотфиксирующие (азотусваивающие) бактерии живут свободно в почве и в воде; другие — в симбиотическом сожительстве с растениями, преимущественно бобовыми. Бактерии поселяются в бородавчатых вздутиях — клубеньках корней этих растений. Отсюда произошло и название этих бактерий — клубеньковые. Необходимую для фиксации азота энергию они получают в процессе окисления безазотистых органических соединений.
Среди свободно живущих азотфиксирующих бактерий наибольшее значение имеют следующие: аэробные бактерии рода азотобактер (Аzоtоbасtег) — слегка приплюснутые кокки, часто объединенные попарно, имеющие слизистую капсулу; анаэробные бактерии, открытые С. Н. Виноградскнм, Clostridium pasterianum — подвижные спорообразующие палочки, способные сбраживать углеводы по типу масляно - кислого брожения, которое и служит этим бактериям источником энергии для связывания молекулярного азота.
Азотфиксирующие бактерии имеют большое значение. В результате их деятельности почва обогащается доступной для растений формой азотистого питания.
В практике сельского хозяйства в качестве бактериального удобрения используют препараты азотфиксирующих бактерий — азотобактерин (из культур азотобактера) и нитрагин (из культур клубеньковых бактерий).
БИОЛОГИЯ И МИКРОБИОЛОГИЯ
РАЗДЕЛ 1 БИОЛОГИЯ
Лекция № 1 Современный этап развития биологии
Дата: 2019-11-01, просмотров: 428.