Агентов
Сравнение антибактериальной активности фторхинолонов с 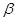 -лактамными (пенициллинами, цефалоспоринами) и другими антибиотиками свело эти две совершенно разные по своему происхождению группы антибактериальных агентов в рассмотрение в одни и тех же книгах [8]. Следует подчеркнуть, однако, что фторхинолоны не имеют отношения к биотехнологии и являются продуктами синтетическими. Это отличает их от полусинтетических антибиотиков, отдельные фрагменты которых получают с участием микроорганизмов [5].
-лактамными (пенициллинами, цефалоспоринами) и другими антибиотиками свело эти две совершенно разные по своему происхождению группы антибактериальных агентов в рассмотрение в одни и тех же книгах [8]. Следует подчеркнуть, однако, что фторхинолоны не имеют отношения к биотехнологии и являются продуктами синтетическими. Это отличает их от полусинтетических антибиотиков, отдельные фрагменты которых получают с участием микроорганизмов [5].
Различен и механизм действия фторхинолонов. Соединения этого класса обладают способностью проникать через клеточные мембраны и селективно воздействовать на размножение бактерий путем ингибирования бактериальной ДНК-топоизомеразы II (ДНК-гиразы) – фермента, отвечающего за разрыв и восстановление суперскрученной спирали ДНК. Согласно современным представлениям, молекулы хинолона связываются с ДНК, образуя сложный комплекс из четырех молекул фторхинолона, двойной спирали ДНК- и ДНК-гиразы [6]. Образование такого комплекса происходит в тот момент, когда гираза осуществляет разрезание обеих цепей ДНК. Тем самым нарушается процесс размножения бактерий. Следует отметить, что механизм действия фторхинолонов принципиально отличается от механизмов действия других групп бактериальных препаратов: пенициллинов, цефалоспоринов, аминогликозидов, сульфамидов, что особенно важно для лечения инфекционных заболеваний, вызванных резистентными к этим препаратам штаммами бактерий [8].
Механизм действия фторхинолонов
Все фторхинолоны действуют бактерицидно благодаря ингибированию ферментов класса топоизомераз – ДНК-гиразы (топоизомеразы II) и топоизомеразы IV [16].
Эти ферменты выполняют строго определенные функции в процессе формирования пространственной структуры молекулы ДНК при ее репликации: ДНК-гираза катализирует расплетение (отрицательиую суперспирализацию) нитей ДНК, а топоизомераза IV участвует в разъединении (декатенации) ковалентно-замкнутых кольцевых молекул ДНК. Ингибированне этих ферментов нарушает процессы роста и деления бактериальной клетки, что приводит к ее гибели [23].
Основной мишеныо в грамположительных микроорганизмах преимущественно является топоизомераза IV, а в грамотрицательных – ДНК-гираза [6].
Механизмы резистентности
Развитие резистентности связано с мутациями в генах gyrA и gyrB (кодируют ДНК-гиразу), ра r С ( grlA ) и _ are ( grlB ) – кодируют топоизомеразу IV, а также в гене nor А (кодирует мембранные белки, которые участвуют в активном выбросе – эффлюксе – фторхинолонов из клетки). Высокий уровень резистентности возникает вследствие сочетания этих механизмов [7].
Мутации, возникающие в генах gyrA , gyrB , ра r С и ра r Е, достаточно значительно влияют на активность фторхинолонов I, II, III поколений, а на активность моксифлоксацина – фторхинолона IV поколения – меньше. Например, мутации у S . Aureus в генах, кодирующих топоизомеразы, меньше снижают активность моксифлоксацина, чем ципрофлоксацина, офлоксацина, левофлоксацина, спарфлоксацина [8].
У Escherichia coli двойная мутация гена gyrA приводит к снижению IC150 норфлоксацниа, ципрофлоксацина и спарфлоксацина по сравнению с таковой у немутировавшего типа более чем в 500 раз, в то время как для моксифлоксацина этот показатель не превышает 12 раз [5].
Эффлюкс (мутация в гене nor А) значительно меньше влияет на активность гидрофобных препаратов, таких, как моксифлоксацин, по сравнению с таковой у гидрофильных препаратов, например у ципрофлоксацина.
При применении моксифлоксацина вероятность развития резистентности у грамположительных микроорганизмов, возможно, ниже, чем при применении других фторхинолонов, что связано с его высоким сродством как к топоизомеразе IV, так и к ДНК-гиразе [6].
Дата: 2019-07-30, просмотров: 350.