Лишайники – очень интересная и своеобразная группа низших растений.
В лишайнике сочетаются два организма с противоположными свойствами: водоросль (чаще зеленая), которая в процессе фотосинтеза создает органическое вещество, и гриб, потребляющий это вещество. Взаимоотношения их строятся следующим образом. Гриб получает от водоросли органические вещества – углеводы, но в то же время как бы предоставляет водоросли, находящейся внутри тела лишайника, среду обитания, защиту от пересыхания и перегревания и т. д. Гриб снабжает водоросль достаточным количеством воды и растворенных в ней минеральных солей, которые он сам поглощает из окружающей среды (субстрата, атмосферного воздуха). Таким образом, хотя гриб в некоторой степени паразитирует на водоросли, но и она извлекает из совместной жизни с ним определенную пользу. Следовательно, в этом сожительстве наряду с паразитизмом имеются и черты симбиоза. В результате этих сложных взаимоотношений в процессе эволюции возник новый самостоятельный комплексный организм, имеющий новые, только ему присущие особенности строения и образа жизни и свои закономерности развития. При этом водоросль, выделенная из лишайника, в благоприятных условиях чаще всего может и вне его расти и развиваться самостоятельно, гриб же в этом случае обычно быстро погибает, так как в процессе приспособления к совместному существованию почти полностью потерял способность к самостоятельному развитию.
Поскольку основным формообразующим компонентом лишайников является гриб, то в последнее десятилетие среди ботаников получила распространение точка зрения, рассматривающая лишайники не как самостоятельную группу растений, а как грибы, включающие в свое тело водоросли, – так называемые лихенизированные грибы, которые перешли к паразитизму на водорослях и в связи с этим значительно изменились. При таком подходе виды лишайников распределяются среди грибов в соответствии с типом плодового тела лишайникового гриба и особенностями его строения. Однако своеобразная форма тела лишайников, присутствие особых веществ, неизвестных ни у грибов, ни у водорослей, особенности обмена веществ и ряд других признаков позволяют рассматривать лишайники как самостоятельную группу организмов [4].
1.4 Строение лишайников
1.4.1 Внешнее строение таллома
По форме и величине лишайники разнообразны, их размеры – от нескольких до десятков сантиметров. Вегетативное тело лишайников представлено слоевищем и в зависимости от образующегося в них пигмента может быть серым, сизым, зеленоватым, буро-коричневым, желтым, оранжевым или почти черным [5]. Пигменты лишайников откладываются в оболочках грибных гиф, реже в протоплазме. На интенсивность окраски сильное влияние оказывает освещенность: чем больше освещенность, тем ярче окраска.
Наиболее яркая окраска у лишайников высокогорных и приполярных районов Арктики и Антарктики. Это связано с тем, что пигменты, словно светофильтры, защищают лишайники от жесткого ультрафиолета, а в приполярных районах темноокрашенные лишайники быстрее нагреваются, снег и иней на их поверхности тает и превращается в капельножидкую влагу, необходимую лишайникам [6].
По внешнему виду различают три типа талломов лишайников: накипные (или корковые), листоватые и кустистые; эти типы связаны между собой переходными формами. Кроме них нередко различают еще чешуйчатый и филаментозный (нитевидный) типы талломов [7].
Таллом накипных лишайников представляет собой корочку, очень прочно срастающуюся с субстратом – корой деревьев, обнаженной древесиной, поверхностью скал и камней. Этот таллом невозможно отделить от субстрата, на котором он растет, не повредив его. Поверхность такого накипного таллома может быть порошковатой, зернистой, бугорчатой или реже гладкой; окраска – различной, обычно неяркой. У некоторых накипных лишайников почти весь таллом врастает в субстрат и на поверхности видны только плодовые тела лишайникового гриба; чаще это блюдцевидные апотеции.
Листоватые лишайники имеют вид чешуек или довольно крупных пластинок. Их таллом прикрепляется к субстрату обычно на большей своей части с помощью пучков грибных нитей (гиф) – ризин или отдельных тонких гиф – ризоидов. Лишь у немногих листоватых лишайников таллом срастается с субстратом только в одном месте с помощью мощного пучка грибных гиф, называемого гомфом. Между накипными и листоватыми лишайниками есть промежуточные формы, у которых таллом в центре накипной, а по краю листоватый, отстающий от субстрата.
У кустистых лишайников таллом состоит из ветвей или более толстых, часто ветвящихся стволиков. Такой кустистый лишайник срастается с субстратом только своим гомфом и растет вертикально либо наискось вверх (напочвенные виды) или свисая вниз (виды, растущие на стволах и ветвях деревьев). Между кустистыми и листоватыми талломами могут быть и переходные формы.
Для некоторых кустистых лишайников характерен так называемый первичный таллом, накипный или чаще листоватый, состоящий из мелких чешуек. На первичном талломе уже развивается вторичный, собственно кустистый, таллом в виде отдельных неразветвленных или разветвленных, вертикально стоящих веточек (рис. 4). Эти ветви вторичного таллома называются подециями и особенно характерны для рода кладония (С1аdоniа Hill. em. Vain.), у которого они очень разнообразны по форме. Подеции могут быть палочковидные, шиловидные, туповатые вверху или расширенные в виде бокальчика (сцифовидные), в виде кустиков с обильно разветвленными ветвями (рис. 4, 2). Сцифовидиые подеции часто пролиферируют, т. е. из центра сцифы или по ее краям вырастают новые сцифовидные подеции. У лишайников из родов кладония и стереокаулон (Stеrеосaulon Schreb.) на подециях развиваются чешуйки – филлокладии. Они соответствуют по происхождению чешуйкам первичного таллома, как бы переходящим на подеции (рис. 4, 3). Первичный таллом может сохраняться в течение всей жизни лишайника, но у ряда видов он быстро исчезает и остаются только подеции.
Талломы большинства кустистых лишайников имеют радиальное строение (у кладонии, стереокаулона, уснеи, алектории). Есть кустистые лишайники с лентовидным ветвящимся талломом, где морфологически хорошо различаются две стороны – верхняя и нижняя (так называемое дорзовентральное строение). Такой таллом характерен для рода цетрария (Сеtraria Ach.) [4].

Рисунок 4 – Таллом кладоний: 1 – таллом (а – первичный, б – вторичный, или подеций), 2 – подеции кладоний (а, б – шиловидные, или палочковидные, в – с апотециями, г – сцифовидный с пролиферацией по краю, е – сцифовидный с пролиферацией в центре, ж – кустистый), 3 – талломы с филлокладиями, 4 – филлокладии [а – дланевидно(пальчато) разветвленные, б – бородавчатые (бугорчатые), в – коралловидно разветвленные, г – щитовидно-чешуйчатые].
1.4.2 Грибы и водоросли, входящие в состав таллома лишайников
Среди лихенизированных грибов 90% связаны с зелеными водорослями, способными фиксировать атмосферный азот, а остальные 10% - с сине-зелеными водорослями из родов Nostoc Adanson, Scytonema Ag., Stigonema Ag., Ehichothrix Ag. и Calothrix Ag., усваивающими азот из атмосферы. Удивительно, что эти лишайники не характерны для местообитаний с недостатком азота, хотя водоросли в составе их таллома, безусловно, его фиксируют и высвобождают в значительном количестве [8].
Из одноклеточных зеленых водорослей в лишайниках встречаются виды родов хлорелла (Сhlоrеllа Beyer), цистококк (Сystoсоссиs Nüg); из нитчатых зеленых водорослей – кладофора (С1аdорhоrа Kütz), плеврококк (Рleuroсоссus auct.), трептеполия (Тrеntероhliа Mart.). При этом нити плеврококка и трентеполии в талломе лишайника часто распадаются на отдельные клетки. Виды этих родов – обычные, широко распространенные водоросли, часто встречающиеся как свободноживущие в пресных водоемах или па стволах деревьев. Нитчатая зеленая водоросль требуксия (Тrеbоихiа Puym.) живет только в талломах лишайников. Из сине-зеленых водорослей в талломах лишайников чаще всего встречаются представители рода носток (Nostoс Adanson); нитчатое тело этих водорослей в талломе лишайника также обычно распадается на отдельные участки. Водоросль, входящая в состав таллома лишайника, называется фотобионт.
Грибы, входящие в состав лишайников, в основном относятся к классу сумчатых (Аsсотусеtеs). Гифы гриба в подавляющем большинстве случаев (за исключением слизистых лишайников) составляют основу таллома лишайников, водоросль же как бы заключена между гифами гриба. Гриб, входящий в состав таллома лишайника, называется микобионт [4]
1.4.3 Анатомическое строение таллома
По анатомическому строению различают два типа талломов лишайников. В более примитивном, гомеомерном талломе клетки или нити водоросли более или менее равномерно распределены между гифами гриба по всей толщине таллома. Такой таллом имеют слизистые лишайники, содержащие в качестве фикобионта нитчатые сине-зеленые водоросли. Нити этих водорослей окружены большим количеством слизи, в которой по всем направлениям проходят грибные гифы, что хорошо видно на поперечном срезе таллома под микроскопом или через 10-кратную лупу. В сухом состоянии такие талломы имеют вид буроватых или черноватых морщинистых корочек. Во влажную погоду они быстро впитывают влагу, разбухают и приобретают форму листоватых лишайников [9]. Один из таких лишайников – коллема (Соllета Web.), виды которого широко распространены на скалах и камнях на Черноморском побережье Крыма, Кавказа и сопредельных областей. Это черноватые или темно-оливковые подушечки, образуемые извилистыми приподнятыми лопастями.
Более сложно построен гетеромерный таллом. На поперечном срезе такого таллома под микроскопом хорошо различимы элементы его строения. Верхняя кора таллома образована плотным переплетением гиф гриба. Дальше идет гонидиальный слой, состоящий из клеток водоросли. Гифы гриба, заходящие в гонидиальный слой, образуют мелкие разветвления, которые плотно примыкают к клеткам водоросли. Здесь гриб получает от фотосинтезирующей водоросли углеводы. Дальше расположена сердцевина (слой из рыхлопереплетенпых гиф гриба), с помощью, которой внутри таллома поддерживаются определенная влажность и воздушная среда, необходимая и для самих гиф, и для клеток водоросли. За сердцевиной расположена нижняя кора из плотно переплетенных гиф гриба. Гетеромерный таллом хорошо выражен у листоватых лишайников и у кустистых лишайников с лентовидным дорзовентральным талломом.
У кустистых лишайников с цилиндрическим радиальным талломом (сцифовидный, палочковидный и т. д.) таллом имеет гетеромерно-радиальное строение. Под корой, одевающей снаружи ветви такого таллома, лежит гонидиальный слой, идущий вокруг всего таллома, а внутрь от него расположена сердцевина. У накипных лишайников с гетеромерным талломом никогда не бывает нижней коры, они срастаются с субстратом сердцевинными гифами.
Развитие и выраженность слоев гетеромерного таллома у разных видов лишайников различны. У ряда листоватых и кустистых лишайников с гетеромерным талломом происходит местный разрыв корового слоя и возникают плоские, неотчетливо очерченные пятна, обычно более светлые, чем сама кора (макулы, или цифеллы); они служат для проведения воздуха в сердцевину таллома и у ряда видов являются определительным систематическим признаком. Встречаются макулы у рода цетрария, например у «исландского мха» (Сеtraria islandicа L.)
1.4.4 Органы спороношения лишайникового гриба
На талломе лишайника из грибных гиф формируются плодовые тела гриба со спорами. Это в основном расположенные на поверхности таллома апотеций или погруженные в таллом кувшиновидные перитеции.
В апотециях и перитециях формируются споры для размножения лишайникового гриба.
Перитеции встречаются у небольшого числа лишайников. Апотеции образуются у многих видов лишайников. Они имеют чаще блюдцевидную форму, но встречаются и более или менее выпуклые или почти шаровидные. Самые крупные апотеции имеют диаметр более 1 см, но лишайников с такими апотециями немного. У громадного большинства лишайников диаметр апотециев от одного до нескольких миллиметров. Они разбросаны по поверхности листоватого таллома, чаще в его середине, или расположены по краям его лопастей. У кустистых лишайников апотеции обычно находятся на концах веточек или сциф. Они бывают сидячими или реже на небольших ножках приподнимаются над талломом. Апотеции могут быть окрашены в один цвет с талломом, или поверхность апотеция (так называемый диск апотеция) имеет другую окраску.
По строению различают три типа апотециев: леканориновый, лецидеиновый и биаториновый.
Леканориновый апотеций по своему анатомическому строению сходен с талломом лишайника. Его диск имеет талломный (слоевищный) край, образованный талломом лишайника и состоящий из гиф гриба и клеток водоросли. Этот край окрашен так же, как и таллом лишайника, и отличается по окраске от самого диска. В самом талломе под леканориновый апотецием также имеются клетки водоросли (рис. 5, 2).
Лецидеиновый апотеций имеет край, состоящий только из гиф гриба и окрашенный в тот же цвет, что и диск апотеция. В этом крае, а также в талломе под лецидеиновым апотецием клеток водорослей нет. Лецидеиновые апотеции имеют твердую консистенцию и обычно темную окраску (рис. 5, 3).
Биаториновый апотеций имеет такое же строение, как лецидеиновый, но отличается яркой окраской и мягкой консистенцией.
Размножение лишайников
Споры лишайникового гриба, выбрасываемые из перитециев и апотециев, прорастают в гифы, которые, переплетаясь, образуют зачаточный таллом лишайника, или прототаллом. Чтобы в дальнейшем из него возник настоящий лишайник, необходим контакт прототаллома с определенным видом водоросли, а также определенные внешние условия. В противном случае прототаллом быстро погибает. Таким образом, споры гриба, образующиеся в апотециях и перитециях, большого значения для размножения лишайников не имеют, поскольку сочетание необходимых для этого условий в природе встречается редко.
Более важны для размножения лишайников такие образования, в которых одновременно присутствуют гифы гриба и клетки водоросли. Это соредии и изидии. Они служат для размножения лишайника как целого организма. Попав в благоприятные условия, они дают начало непосредственно новому таллому. Соредии и изидии встречаются чаще у листоватых и кустистых лишайников.
Соредии представляют собой мельчайшие образования в виде пылинок, состоящих из одной или нескольких клеток водоросли, окруженных гифами гриба (рис. 5, 4). Формирование их обычно начинается в гонидиальном слое. Вследствие массового образования соредии количество их увеличивается, они давят на верхнюю кору, разрывают ее и оказываются на поверхности таллома, откуда легко сдуваются при любом движении воздуха или смываются водой. Скопления соредии называют соралями. Наличие и отсутствие соредии и соралей, их расположение, форма и окраска постоянны для определенных лишайников и служат определительным признаком.
Иногда при отмирании лишайников их таллом превращается в порошковатую массу, состоящую из соредии. Это так называемые лепрозные формы лишайников (от греческого слова «лепрос» — «шероховатый», «неровный»). В этом случае определить лишайник почти не представляется возможным.
Соредии, разносимые ветром и дождевой водой, попав в благоприятные условия, постепенно образуют новый таллом. Возобновление нового таллома из соредии происходит очень медленно. Так, у видов из рода кладония нормальные чешуйки первичного таллома развиваются из соредии только через срок от 9 до 24 месяцев. А для развития вторичного таллома с апотециями требуется от одного до восьми лет в зависимости от вида лишайника и внешних условий.
Если условия для развития нового таллома не совсем благоприятны, но все же возможен рост грибных гиф и размножение водорослей, то из соредии развиваются новые соредии и возникает описанная выше лепрозная форма, имеющая вид порошковатого пятна.
Изидии встречаются у меньшего числа видов лишайников, нежели соредии и сорали. Они представляют собой простые или коралловидно разветвленные выросты, обычно густо покрывающие верхнюю сторону таллома. В отличие от соралей изидии снаружи покрыты корой, часто более темной, чем таллом. Внутри, под корой, они содержат водоросли и грибные гифы (рис. 5, 5). Изидии легко отламываются от поверхности таллома. Обламываясь и распространяясь с помощью дождя и ветра, они, так же как и соредии, могут при благоприятных условиях образовывать новые талломы лишайников.
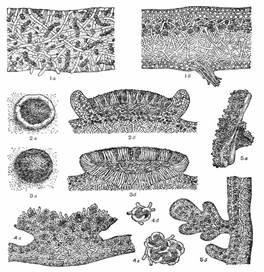
Рисунок 5 – 1 – разрез таллома (а – гомеомерного, б – гетеромерного) 2 – леканориновый апотеций (а – вид сверху, б – разрез), 3 – лецидеиновый апотеций (а – вид сверху, б – разрез), 4 – строение соралей (а – сораль, б, в – отдельные соредии), 5 – изидии (а – внешний вид, б – разрез).
Многие лишайники не образуют апотециев, соредий и изидии и размножаются участками таллома, которые легко отламываются от хрупких в сухую погоду лишайников ветром или животными и ими же переносятся. Особенно широко распространено размножение лишайников участками таллома в арктических областях. В наибольшей степени размножение кусочками таллома присуще накипным эпилитным лишайникам. Появление трещин в скалах и камнях, покрытых лишайниками, а также появление трещин в самом талломе накипного лишайника из-за неодинакового отношения к нагреванию и увлажнению камня и самого лишайника ведет к разламыванию таллома и отделению небольших его участков. Эти отделившиеся кусочки таллома и переносятся ветром и водой на новые места, давая начало новым талломам.
Для некоторых лишайников (около 100 видов из разных родов) характерно наличие на поверхности, а иногда и внутри таллома образований разного размера в виде разбросанных небольших вздутий, бородавочек, булавовидных, шаровидных и коралловидных выростов. Это цефалодии. Они могут покрывать верхнюю или нижнюю сторону листоватых талломов или развиваться по бокам подециев либо веточек кустистых талломов. В отличие от изидии они более разнообразной формы и разбросаны более редко. На срезе под микроскопом видно, что они имеют гомеомерное строение и состоят из беспорядочного скопления гиф, среди которых равномерно распределены клетки или нити сине-зеленых водорослей. Реже цефалодии имеет гетеромерное строение. Водоросли в цефалодиях всегда сине-зеленые, тогда как в талломе лишайника, несущего цефалодии, водоросли всегда зеленые. Образование цефалодиев обусловлено проникновением в таллом не свойственных данному виду лишайника сине-зеленых водорослей, которые вызывают местное разрастание гиф гриба, «обрастающего» эти водоросли. Роль цефалодиев для самого лишайника не совсем ясна. Предполагается и отчасти доказано экспериментально, что они увеличивают питание лишайника, живущего на бедном субстрате. Сине-зеленые водоросли цефалодия способны фиксировать атмосферный азот, который затем в виде азотистых соединений усваивается всем талломом лишайника – составляющими его грибом и зеленой водорослью, которые сами не могут фиксировать атмосферный азот.
Цефалодии специфичны для ряда видов и имеют значение определительного (систематического) признака. Характерный пример лишайника с цефалодиями – напочвенный вид пельтигера пупырчатая ( Peltigera aphtnosa (L.) Willd.).
Дата: 2019-07-30, просмотров: 476.