Важкі метали є протоплазматичними отрутами, токсичність яких збільшується по мірі збільшення відносної атомної маси.
За фітотоксичністю сполуки можна розподілити у такий ряд:
• дуже фітотоксичні елементи – чинять дію на тест-організми при концентраціях в розчині до 1 мг/л: Аg+, Ве2+, Нg2+, Sn2+ і можливо Со2+, Ni2+, Pb2+ і СrO2- .
• помірно токсичні: 1-100мг/л викликає інгібуючу дію: AsO42-, ВO33-, ВгО4-, СІО4-, МnО42-, МоО42- антимоніт селену; іони As, Se, Al, Ba, Cd, Cr, Fe, Mn, Zn.
• слаботоксичні: рідко чинять негативний ефект при рівнях більше 1800 мг/л: Сl-, Вr-, L-, Са2+, Mg2+, K+, Na+, Rb2+, Sr2-, Li+ , NO3- .
Токсичність важких металів може проявлятися по-різному:
- Сu, Нg: при токсичній концентрацій інгібують активність ферментів, утворюють комплекси з органічними молекулами, що здатні проникати крізь клітинну мембрану.
- Al, Ba, Fe: утворюють преципітати з РО43-, SO42-, утворюють хелатоподібні комплекси з обмінними метаболітами.
Взаємодіють із клітинними мембранами, змінюючи їх проникність і інші властивості. Au, Cd, Cu і Fe2+ іноді викликають розрив клітинних мембран.
Конкуренція: Li – з Na; Cs – з К; Ва і Sr – з Са; Cd – з Zn.
На фітотоксичність впливають такі показники: концентрація металу в ґрунтовому розчині. Деякі рослини здатні акумулювати метали; ґрунтові фактори: рН (впливає на рухомість металів і засвоєння їх кореневою системою), вміст органічної речовини та інші фактори. При рН близько 7 у ґрунтах з істотним вмістом важких металів попереджується фітотоксичність багатьох з них, але ті ж концентрації при рН 5,5 і нижче можуть стати летальними для рослин; забезпеченість рослин елементами живлення, фази росту, глибини проникнення коренів, тривалості вегетаційного періоду; удобрення, вапнування; зміна освітленості, температури і зволоження впливає на рух і трансформацію важких металів у ґрунтовому середовищі і рослинах, на взаємодію між рослинами і металами.
Для характеристики вибіркового поглинання хімічних елементів рослинами Б.Б. Полинов (1945) ввів у обіг величину, що пізніше була названа А.І. Петельманом (1961) коефіцієнтом біологічного поглинання (КБП), що представляє собою часткове відношення кількості хімічного елемента у золі рослин до його вмісту в літосфері [16, 105]:
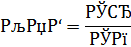
де Ср – вміст елемента в золі рослини; Сп – вміст елемента в ґрунті.
КБП змінюється від 0,001 до 100. У складених ним рядах біологічного поглинання виділено п'ять груп елементів: інтенсивно накопичувані – КБП=10...100; сильно накопичувані – КБП=1...10; слабо накопичувані і середнього захвату – КБП=0,1...1,0; слабкого захвату – КБП=0,001-0,01. КБП хімічного елемента для одного і того ж виду рослин не є сталим і може змінюватись.
Сучасні уявлення фізіологів щодо надходження і поведінки важких металів у рослинах (Вахмістров, 1966; Tiffin, 1977; Вахмістров, Мазель, 1973; Лібберт, 1976; Клаксон, 1978) зводяться до наступного [16, 116].
Важливу роль у захисті рослин від надлишку важких металів, що надходять із ґрунту в корені, виконує коренева система. Затримуючи надлишкові іони, корені тим самим сприяють зберіганню в надземних органах сприятливих (чи нешкідливих) концентрацій хімічних елементів.
Захисні можливості коренів дуже великі, але і вони мають межу; при дуже високій кількості токсиканта у середовищі його потік у надземні органи посилюється. Спочатку це відмічається у листках, потім – і у зернах.
Іони металу, потрапивши у корінь, займають вільний простір, адсорбуються на стінках і залишаються в розчині. Щоб брати участь у метаболізмі коренів, їм необхідно подолати плазмалему. Подолання клітинних мембран необхідне і для досягнення іонами ксилеми: обійти перешкоду – поясок Каспарі – вони можуть тільки шляхом переходу із апопласта у симпласт. Однак цей шлях долається важко, оскільки у мембранах локалізований механізм вибіркового поглинання іонів, який обмежує проникнення у клітину баластних і надлишкових іонів. Якщо все ж таки у клітинах кореня іонів виявиться понад допустимий ліміт, то включається ще один механізм захисту, який переводить надлишок у вакуолі.
Таким чином, частина іонів металу затримується у вільному просторі чи переправляється у вакуолі, інша частина використовується у процесі метаболізму, третя – із ксилемним соком підіймається у надземні органи.
При просуванні по ксилемі метали можуть адсорбційно поглинатися її стінками, а також закомплексуватися присутніми у клітинному соці органічними сполуками. Однак, як вказує У. Ліндсей (Lindsay, 1972), важкі метали – Zn, Ni, Cu, Fe – не зв'язані тут у високостабільні ліганди [16, 123]. Проходячи переважно транзитом шлях по ксилемі, іони потрапляють у листки, перш за все, в апопласта. Для того, щоб проникнути в клітини листка, у якому відбувається основна синтетична діяльність рослин, іонам знову потрібно подолати клітинні мембрани. За аналогією з коренями тут діють механізми вибіркового поглинання. Основна його функція – забезпечення нормального вмісту іонів у цитоплазмі, тобто вона пов'язана з захистом життєво важливих органів і процесів. При потраплянні важких металів у листок їх надлишок може акумулюватися у різноманітних структурах листка і клітини: провідні тканини, апопласта, вакуолі.
Як відомо, елементи з ґрунту поглинаються переважно у вигляді іонів. Однак у рослинній тканині ця форма вже не домінує. Метали в іонній формі, очевидно, у помітній кількості можуть зустрічатися в ксилемі, апопласті і вакуолях, тоді як у цитоплазмі вони входять, в основному, у склад органічних сполук. Органічні сполуки, що виходять за межі клітин, можуть хелатувати іони металів, роблячи їх менш активними і пом'якшуючи тим самим вплив несприятливого ефекту (Wallace et. c, 1968) [16, 127].
За ступенем рухомості А.Л. Курсанов (1976) виділяє у тканинах три зони [16, 132]:
- вільний простір, звідки іони вимиваються легко;
- цитоплазму, яка важко віддає іони;
- вакуолі, де іони утримуються міцно.
Викладені уявлення допомагають пояснити ситуації, що виникають при забрудненні середовища важкими металами.
Важкі метали негативно впливають на різні боки життя рослин. М.Д. Степановою і Н.Ю. Гармаш було встановлено, що при високому вмісті у рослинних тканинах свинцю змінюється кількість N, Р, Са і Мо, при надлишку Cd – Са, Mn, Cu, Мо. Більшою мірою це стосувалося листків, меншою – насіння [19, 231].
При більш детальному вивченні білкового обміну виявилося, що надлишкова концентрація важких металів у рослинах впливає на його фракційний склад, при постійності вмісту глобулінів зростає кількість проламінів, глютелінів і нерозчинного залишку і дещо зменшується вміст альбумінів. Ця обставина вказує на погіршення складу білків, зокрема на зниження вмісту лізину.
Важкі метали, починаючи з певної концентрації, гальмують процес фотосинтезу і зменшують транспірацію рослин. При високому вмісті у середовищі, наприклад РЬ, продуктивність фотосинтезу в різних сільськогосподарських культурах виходить на рівень 10% від максимальної, а транспірація скорочується майже у 20 разів порівняно з контролем (Bazzaz et al., 1985) [16, 148].
Результатом пригнічення токсикантами фізіолого-біохімічних процесів є загальне послаблення опору рослин до хвороб і шкідників, що називається «вторинною» дією токсикантів.
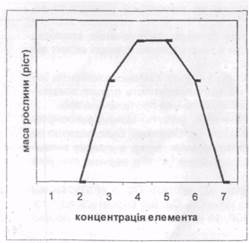
Рис. 1. Фітотоксичність важких металів
Токсичну дію металів на рослини можна прослідкувати по росту:
· 0-2 – ріст відсутній;
· 2-3 – гостра нестача;
· 3-4 – середня нестача;
· 4-5 – оптимальний вміст;
· 5-6 – слабка токсичність;
· 6-7 – сильна токсичність;
· 7 – загибель рослини.
На відміну від симптомів нестачі, які для кожного елементу є специфічними, ознаки надлишку більш-менш однакові. Згідно зі схемою Busser, при поступовому зростанні концентрації іонів у середовищі спостерігається поступова поява ознак пригнічення рослинного організму:
· 1 – гальмування росту;
· 2 – хлороз листків;
· 3 – некрози верхівок і країв листків;
· 4 – відмирання коренів.
Якщо некрози листків і відмирання коренів виступають як прямий наслідок надлишкового вмісту елементу в рослинних тканинах, то хлороз і обмеження росту можуть бути також і результатом антагоністичних взаємовідносин надлишкового іону з іонами поживних речовин і виникнення таким чином індукованої нестачі у тканині.
Висока фітотоксичність властива Нg і Cd. Менш токсичними є Cu, Zn, Pb. У дослідах із зеленними культурами встановлено наступний ряд токсичності вивчених хімічних елементів: Cd > Ni > Zn, Cr > Pb [Foroughi et al., 1975] [19, 239]. Згідно з даними, фітотоксичність важких металів розміщується таким чином: Cd > Cu > Co = Ni > As = Cr > Zn > Mn = Fe > Pb.
K.V. Smilde (1981) розташував метали за фітотоксичністю в такий ряд: Cd> Ni> Cu> Zn> Cr і РЬ. Він вказує на те, що метали у чистому вигляді менш токсичні, ніж у поєднанні з іншими металами.
За чутливістю до кадмію рослини можна розмістити у такій послідовності (по зростаючій): томат, овес, салат, лугові трави, морква, редька, квасоля, горох. Цинк слаботоксичний для рослин, малотоксичний і молібден, навіть якщо він попадає у ґрунт у великій кількості. Мідь у високих концентраціях може мати токсичну дію на рослину, особливо на легких і малогумусних ґрунтах. Ознаки хлорозу й утворення численних зафарбованих у коричневий колір бічних корінців відмічалось у рослин при вмісті у ґрунті 0,7-1,1 кг/га сполук міді, які вилучаютьсь з водою. Найменшу безпеку становить свинець, оскільки у рослинах добре відлагоджена система захисту при проникненні його у кореневу систему.
Дата: 2019-05-28, просмотров: 373.