Возвращаемся к современности. На следующем рисунке показано дерево 37-маркерных гаплотипов гаплогруппы Q наших современников, и в подписи описаны основные ветви гаплогруппы.
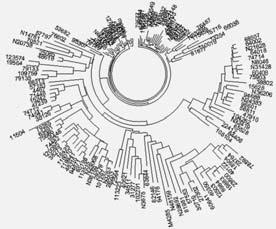
Дерево гаплогруппы О из 140 гаплотипов в 37-маркерном формате. Верхняя компактная ветвь из 37 гаплотипов – группа евреев гаплогруппы О, возрастом 995 ± 110 лет до общего предка. Ветвь справа – европейцы, в основном англичане, шведы и норвежцы, которые имеют древнюю гаплогруппу Q-M236, но следующий нисходящий субклад (L213) у них уже отсутствует. Их общий предок жил 3400 ± 580 лет назад. Этот нисходящий субклад присутствует у другой группы европейцев (на 8 часов на диаграмме выше), среди которых предобладают шведы, англичане, шотландцы, норвежцы, их общий предок жил всего 2100 ± 340 лет назад. Нижняя ветвь – в основном американские индейцы субклада Q-M3. Построено по данным[136].
В сети есть «еврейский проект гаплогруппы Q»[137], на основании которого ветвь наверху дерева на рисунке выше может быть дополнительно развернута, и соответствующее дерево приведено на следующем рисунке.
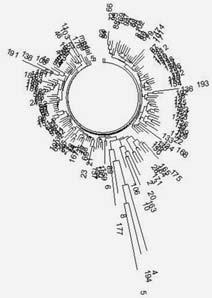
Дерево из 143 гаплотипов ветви евреев гаплогруппы Q в 37-маркерном формате. 127 гаплотипов из них (89 %) занимает однородная группа, общий предок которых жил 995 ± 110 лет назад. Предки их жили в Германии, России, Украине, Белоруссии, Литве, Латвии, Польше, Венгрии. Нижняя узкая ветвь – гаплотипы из Швеции, Испании, Англии, Марокко, Саудовской Аравии, и не очень ясно, потомки ли они евреев. Правое крыло этой узкой ветви занимают гаплотипы меннонитов, которых иногда называют «швейцарскими меннонитами». Построено по данным[138]
Обращает на себя внимание, что европейские евреи, ашкенази, несмотря на то, что относятся к разным гаплогруппам, в своем большинстве происходят от общих предков, которые жили примерно тысячу лет назад. Это – евреи гаплогруппы G2b (1030 ± 140 лет назад), франкфуртские евреи гаплогруппы J2 (1025 ± 120 лет назад), еще одна, значительно отличающаяся ДНК-линия евреев (1025 ± 170 лет назад), европейские коэны гаплогруппы J1 (1075 ± 130 лет назад), евреи гаплогруппы R2 (1050 ± 150 лет назад, см. ниже). Понятно, что евреи не возникли в Европе тысячу лет назад. Первое упоминание о них в Европейских хрониках датируется 321 годом нашей эры, когда римский император Константин обязал декретом направить представителей еврейской общины Кёльна в городской совет[139]. После этого в хрониках не было упоминаний о евреях до 9-го века. На самом деле евреи продолжали жить в Кёльне до 1424 года, когда они были изгнаны из города[140]. Об этом же свидетельствуют и археологические данные, показывающие типичные предметы жизни евреев[141].
Естественно, мы пока не знаем, к каким гаплогруппам те евреи относились, но данные ДНК-генеалогии определенно говорят о том, что в состав еврейского сообщества в Европе произошло вливание новых групп евреев гаплогрупп J1, J2, G2a, Q, R2, R1a (см. ниже). Может быть, хазары, прибывшие из черноморско-каспийского региона? Мы пока не знаем, но скоро определенно узнаем. Интересно, но именно тогда же появилось название «ашкеназы». На самом деле «ашкеназы» – древнее слово, и обозначало царство, наряду с царствами араратским и минийским, как впервые появляется в книге пророка Иеремии, в разделе 51:27. Текст гласит – «Поднимите знамя на земле, трубите трубою среди народов, вооружите против него народы, зовите на него царства Араратские, Минийские и Аскеназские, поставьте вождя против него, наведите коней, как страшную саранчу». На кого же это так? Да на Вавилон, на кого же еще? Почему же вдруг это вспомнили в Европе, тысячи лет спустя? Что там так изменилось? Может быть, действительно потому, что подошли хазарские евреи разных гаплогрупп, которые подняли иудаистику в Европе на новый уровень? Мы это тоже пока не знаем.
Несколько слов о американских индейцах гаплогруппы Q1a2a1a1-M3. Дерево их гаплотипов показано на рисунке. Общий предок дерева жил 16300 ± 3300 лет назад.
Интересно, что самые древние предковые гаплотипы дерева имеют вид
13 13 24 10 14 14,
а современные монгольские гаплотипы той же гаплогруппы Q имеют вид, например,
13 12 23 10 14 13
13 12 24 9 14 14
и общий предок тех монгольских гаплотипов жил 13700 ± 3700 лет назад[142]. Это еще раз показывает, уже фактически на количественном уровне, откуда пришли предки современных американских индейцев. Возможно, в этой связке упрятано сходство чукотско-камчатских языков с языками на-дене, одними из многих языков «нативных американцев».
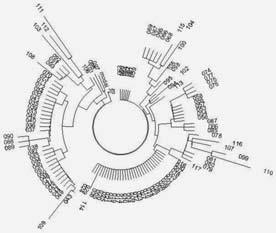
Дерево из 117 гаплотипов американских индейцев субклада Q1a2a1a1-M3 в 6-маркерном формате. Как часто происходит с древними популяциями, деревья их гаплотипов состоят из ряда плоских ветвей, прошедших бутылочные горлышки и недавно выживших. Но сами ветви настолько разбросаны, что из общий предок уходит в глубину времен очень далеко.
Общий предок данного дерева жил 16300 ± 3300 лет назад[143].
Глава 24
Гаплогруппа R
Среди наших современников носители гаплогруппы R* (то есть не считая носителей нижестоящих субкладов) пока не найдены. Это – обычная ситуация для древних гаплогрупп. Вероятность ДНК-линии продержаться без мутаций, нисходящих к мутации десятки тысяч лет назад, крайне мала. Напомним, что мутация во фрагменте Y-хромосомы размером 10 миллионов нуклеотидов происходит, в среднем раз в 144 года, гаплогруппа R образовалась примерно 31300 лет назад, и «под ней» до настоящего времени находятся около двухсот необратимых снип-мутаций. У кого-то их больше, у кого-то меньше, что является отражением статистики мутаций, но так, чтобы у кого-то не проскочило ни одной мутации – таких должно быть очень мало. Хотя наверняка есть, среди десятков миллионов, если не сотен миллионов носителей гаплогруппы R1b с субкладами.
Гаплогруппе R уделяется в исследованиях особое внимание, как предковой гаплогруппе «внуков» – гаплогрупп R1a и R1b, которые преобладают в Восточной и Западной Европе, соответственно.
Поэтому находка древних костных остатков неподалеку от Байкала, рядом с селением Мальта, с археологической датировкой 24 тысячи лет назад, ДНК которых показало гаплогруппу R, стало настоящей мировой сенсацией. Кости принадлежали, как было установлено, 4-летнему мальчику. Автор этой книги предсказывал сибирское происхождение гаплогруппы R1b и ее предков еще в 2012 году, как и последующие миграции гаплогруппы R1b из Сибири (что есть часть Центральной Азии) на запад, до Европы, и об этом даже название статьи[144], но сейчас не об этом речь. Речь о том, что в угаре этой сенсации в средствах массовой информации, особенно на Западе, пошли самые фантастические теории о том, как эта гаплогруппа оказалась в Сибири, и какова ее роль в заселение Америки. На самом деле роли не было никакой, гаплогруппы R среди коренных американцев, то есть американских индейцев, нет, и предки байкальского мальчика или потомки тех предков с территории современной Мальты в Америку не ходили. Ходили носители гаплогруппы Q, а у R и Q – один общий предок, гаплогруппа Р. Поэтому геномы носителей гаплогрупп Q и R, по сути братьев, должны быть весьма похожи. Геномный анализ, понятно, так и показал, что геном мальчика из Мальты на 14–38 % похож на геном современных американских индейцев[145].
Но авторы, как обычно для популяционных генетиков, не посмотрели на гаплогруппы, что у индейцев она преобладающе Q, а у древнего байкальца – R. Потому и валом пошли сообщения, что найдены предки американских индейцев, и это европейцы, потому что именно в Европе находится много гаплогруппы R1b, производной от R. Пошли сообщения, что у русских 13 % генома сходно с американскими индейцами, а значит, там, в древней Америке, генетические корни русских, или напротив, в России генетические корни американских индейцев. А генетики опять не могли сообразить, что никаких корней русских в Америке нет, как и наоборот, просто гаплогруппа Р является предковой как для русских гаплогруппы R1a, так и для индейцев гаплогруппы Q, только и всего. Иначе это как рассуждать так, что я произошел от своего дяди (или наоборот), потому что мой геном похож на геном дяди, и все потому, что у нас общий дедушка.
Но, казалось бы, генетики должны такие случаи различать, или хотя бы об этом думать, и каждый раз выдвигать и обсуждать альтернативные объяснения. Но, к сожалению, популяционные генетики такого не делают, и на гаплогруппы с субкладами не смотрят. То есть они их приводят в статьях (и то не всегда), но в анализ их не включают. Основной принцип, с которым работают популяционные генетики, и который стал полностью определяющим при геномном анализе, является принцип «похожести».
Как это работает, и к чему это приводит? Давайте рассмотрим, это очень важно для понимания, чего стоит современный «геномный анализ» в популяционной генетике.
Главная проблема в геномном анализе – это рассортировать фрагменты ДНК от разных людей так, чтобы отличить закономерные сходства фрагментов от случайных. Закономерные сходства наблюдаются тогда, когда фрагменты ДНК этих людей происходят от недавнего общего предка, где понятие «недавний» определяется поставленной задачей. Такие совпадения называют «идентичные по происхождению» (ИПП). В других случаях совпадения являются случайными, по статистическим причинам. Такие совпадения называют «идентичные по сходству» (ИПС). В категорию ИПС попадают также фрагменты ДНК, которые сходны не случайно, а отвечают за важные функции жизнедеятельности человека, которые сформировались и устоялись в результате эволюции человека. В категорию ИПС попадают также фрагменты ДНК, которые веками циркулируют в рамках определенного этноса, особенно если этнос относительно замкнутый (как, например, в еврейских сообществах прошлого).
Суть геномного анализа состоит в том, чтобы сопоставить ДНК от разных людей (если речь о геномном анализе человека), при том, что ДНК людей похожи буквально на 99 %, и что ДНК испещрены мутациями, то есть заменами одних нуклеотидов на другие, и таких замен в ДНК миллионы. В итоге сравниваются доли процентов различий на фоне более 99 % сходства. Но эти сравнения невозможно производить, пользуясь целыми, интактными молекулами ДНК, они производятся после расщепления ДНК на фрагменты разного размера, и компьютер ищет «матчи», то есть совпадения. «Матчи» имеют разную природу. Одни похожи потому что происходят от одного, сравнительно недавнего общего предка (идентичность по происхождению, ИПП), другие похожи по другим причинам (идентичность по сходству, ИПС), не имеющим никакого отношения к общему происхождению. Или имеющим лишь косвенное отношение, например, потому что отражают близкое этническое происхождение, но по комбинации происхождения по мужской и женской линии, где общего предка нет в принципе. В итоге компьютер, подчиняясь определенным программам, которых в интерпретации ДНК используется множество, сортирует фрагменты по разным критериям одновременно – по числу снипов, по размерам фрагментов, по степени сходства, и ни в одном варианте нет однозначных критериев. Фрагменты бывают «крупные», «менее крупные», «более средние», «менее средние», «умеренно малые», «малые», и всех между ними, а и компьютер сам решает, кто кому родственник «по наследственности», а кто «просто похож».
Всю эту кашу компьютер сортирует, и делает выводы, которые по понятным причинам могут быть совершенно далеки от реальности, или частично пересекаться с реальностью, причем степень этой «частичности» может варьироваться в любых пределах. Понятно, что нужны независимые критерии для перекрестной проверки выводов, но популяционные генетики так не работают. Обычно что получилось, то и получилось. Наиболее очевидным независимым критерием была бы гаплогруппа и субклады, которые тут же вводили запреты на происхождение ДНК одного субклада из другого, но популяционные генетики так тоже не работают. У них критерии простые – совпали протяженные сегменты (фрагменты) ДНК – значит, вероятность ИПП выше, значит, общий предок жил не так давно. Совпадают только малые сегменты – значит, предобладает ИПС. Размеры сегментов измеряют в сантиморганах (сМ), где один сантиморган равен примерно миллиону пар нуклеотидов, или такому расстоянию, на котором рекомбинация генов равна примерно 1 %. Есть и ориентировочные формулы, согласно которым если в двух ДНК в среднем совпадают сегменты размером в и сМ, то там на 99 % ИПП, если совпадают сегменты в 9 сМ, то вероятности ИПП и ИПС – 50 % на 50 %, а если совпадают только сегменты в 4 сМ, то там на 99 % ИПС (случайные совпадения), и всего на 1 % ИПП.
Понятно, что эти критерии весьма условны, как весьма условны и выводы. Остается только удивляться, что иногда выводы разумны, но «разумность» здесь «по понятиям», проверить эту разумность, как правило, не получается. А когда применяется проверка с гаплогруппами-субкладами, вся «разумность», как правило, рассыпается. Но еще чаще бывает, что компьютер в целом описал ситуацию правильно, например, что геномы древнего байкальского мальчика и современных американских индейцев совпадают на 14–38 %, и ясно, что это потому, что и у тех, и у других один общий предок – гаплогруппа Р. Но популяционные генетики делают свою интерпретацию – что популяция того байкальского мальчика перешла в Америку и стала предками американских индейцев, что в корне неверно. Подобные же абсурдные интерпретации делались в перечисленных выше случаях. Поскольку авторы соответствующих работ должны понимать, что все их построения построены на песке, то они излагают свои результаты и выводы в крайне вязком виде, практически никогда не напрямую, но в широкую прессу, в СМИ это уже идет в виде, вполне категоричном. К подобным «выводам» мы еще вернемся, они откровенно дискредитируют науку.
Более того, при подобном «анализе» авторы обычно «подтверждают» то, что уже устоялось в науке, даже когда устоялось неверно. Например, что ямная культура принесла индоевропейские языки в Европу. Это противоречит многим наблюдениям и фактам, но на Западе устоялось (российские историки и лингвисты относятся к этому, как правило, скептически, или отрицательно). Но генетические исследования «подтвердили», об этом речь пойдет в разделе про гаплогруппу R1b. А подтвердили, потому что современная популяционная генетика ничего не оспаривает, если что известно или принято – она всегда подтверждает, только чтобы не подумали, что у них что-то не так.
Вывод – данные «геномного анализа» в популяционной генетике надо непременно проверять перекрестно. Если это пока невозможно, стоит воздержаться от слепого их принятия. Наиболее надежная проверка – с помощью подходов ДНК-генеалогии, сопоставлением гаплогрупп, субкладов, гаплотипов, расчетов датировок.
Помимо «байкальского мальчика», гаплогруппа R была также найдена в Германии в неолитической культуре, с датировкой 5600 ± 60 лет назад, субклад R-P224.
R M207 , P224/PF6050
Глава 25
Гаплогруппа R1
Как и в гаплогруппе R, среди наших современников носители гаплогруппы R1* (то есть не считая носителей нижестоящих субкладов) пока не найдены. Расчетами по количеству снипов, нижестоящих к данной гаплогруппе, показано, что она образовалась примерно 27600 лет назад[146]. Поскольку не найдено ее носителей, то нет и Проекта гаплогруппы R1. Тем не менее, найдены по меньшей мере два древних ископаемых костных остатка носителей гаплогруппы R1, оба в Германии, один с датировкой 4290 ± 85 лет назад, субклад Р234, другой не датирован, субклад Р233, оба найдены в культуре колоколовидных кубков, вместе с носителями гаплогруппы R1b.
• R1 М173/Р241 , M306/PF6147, P225, P231,
P233/PF6142, P234, P236, P238/PF6115, P242/
PF6113, P245/PF6117, P286/PF6136, P294/PF6112
Глава 26
Гаплогруппа R2
Эта гаплогруппа находится в целом на периферии научного интереса западных исследователей, что и находит отражение в степени проработанности ее субкладов:

Дата: 2019-03-05, просмотров: 410.