Основные компартменты клетки:
Клетка — главный гистологический элемент
Три основных компартмента:
· плазматическая мембрана
· ядро
· цитоплазма
Органеллы, включения
Биологические мембраны входят в состав каждого клеточного компартмента и многих органелл.
Компартмент - отдельная, функционально значимая часть клетки или обособленная субклеточная структура, характеризующаяся специфическими морфологическими или биохимическими свойствами и отделённая от остальной клетки по крайней мере одной избирательно проницаемой мембраной.
Химический состав (плазмолемма, клеточная мембрана):
1. Липиды
2. Белки (50%)
3. Углеводы (2–10%)
Функции:
· избирательная проницаемость
· межклеточные взаимодействия
· эндоцитоз
· экзоцитоз
Избирательная проницаемость - поддерживает клеточный гомеостаз, оптимальное содержание в клетке ионов, воды, ферментов и субстратов.
Пути реализации:
• пассивный транспорт
• облегчённая диффузия
• активный транспорт
Ионные каналы (натриевые, калиевые, Ca, хлорные каналы, водные каналы) (аквапорины)
Активный транспорт — энергозависимый трансмембранный перенос против электрохимического градиента, происходящий при участии АТФаз
Натрий, калиевая АТФаза (Na,K-АТФаза) Протонная и калиевая АТФаза (H,K-АТФаза) Са-транспортирующие АТФазы (Ca-АТФаза)
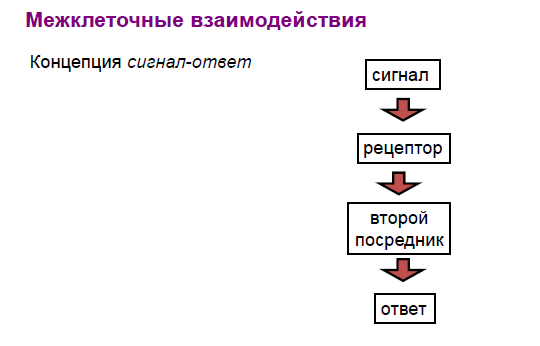
Межклеточные взаимодействия
Плазматическая мембрана — место приложения раздражителей
Из внешней среды:
• физических(например, кванты света в фоторецепторах)
• химических(например, вкусовые и обонятельные молекулы, рН)
• механических(например, давление или растяжение в механорецепторах)
Из внутренней среды организма:
сигналов информационного характера (например, гормоны, нейромедиаторы)
Эндоцитоз — поглощение (интернализация) клеткой веществ, частиц и микроорганизмов.
Варианты эндоцитоза:
· Пиноцитоз — процесс поглощения жидкости и растворённых веществ с образованием небольших пузырьков. Пиноцитозные пузырьки формируются в специализированных областях плазматической мембраны — окаймлённых ямках
· Фагоцитоз — поглощение крупных частиц (например, микроорганизмов или остатков клеток).
· Опосредуемый рецепторами эндоцитоз характеризуется поглощением из внеклеточной жидкости конкретных макромолекул.
·
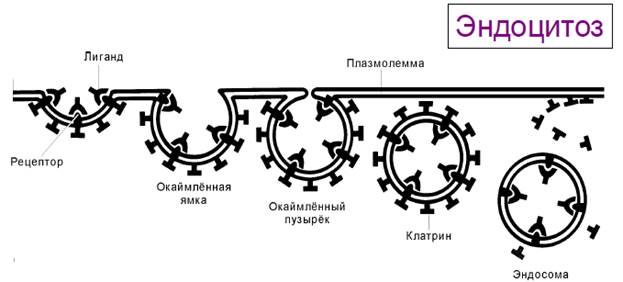
Опосредуемый рецепторами эндоцитоз. Многие внеклеточные макромолекулы (трансферрин, ЛНП, вирусные частицы и др.) связываются со своими рецепторами в плазмолемме. Образуются клатриновые окаймлённые ямки, а затем — окаймлённые пузырьки, содержащие комплекс лиганд-рецептор. Окаймлённые пузырьки после освобождения от клатрина — эндосома. Внутри эндосом лиганд отщепляется от рецептора.
Экзоцитоз — процесс, при котором внутриклеточные секреторные пузырьки (например, синаптические) и секреторные гранулы сливаются с плазмолеммой, а их содержимое освобождается из клетки — секреция .
Спонтанная и регулируемая секреция. Одна часть пузырьков постоянно сливается с клеточной мембраной (спонтанная секреция), в то время как другая часть скапливается под ней, и процесс слияния происходит только под действием сигнала, чаще всего вследствие увеличения концентрации Са2+ в цитозоле (регулируемый экзоцитоз).
1. Спонтанная секреция обеспечивает встраивание в плазмолемму вновь синтезированных белков и рецепторов, интернализованных при опосредованном рецепторами эндоцитозе.
2. Регулируемый экзоцитоз. В этом процессе участвуют секреторные гранулы, а также специализированные эндосомы (например, синаптические пузырьки).
Ядро
23 пары хромосом (диплоидный набор) ядра соматической клетки содержат ядерный геном (примерно 20-30.000 генов).
Реализация генетической информации (транскрипция ® процессинг ® трансляция ® посттрансляционная модификация) и другие функции ядра происходят при участии ДНК и разных видов РНК.
Структурные компоненты ядра: хроматин, ядерная оболочка, ядрышко, нуклеоплазма.
ДНК состоит из двух полинуклеотидных цепей, образованных нуклеотидами, соединёнными фосфодиэфирными связями (1). Ковалентные фосфодиэфирные связи соединяют 5’- атом углерода одного нуклеотида с 3’- атомом углерода следующего нуклеотида. Антипараллельные цепи комплементарно спарены (2): аденин (A) с тимином (T), гуанин (G) c цитозином (C). Клетки перед каждым делением воспроизводят (реплицируют) ДНК (3): дочерние молекулы ДНК воспроизводятся при помощи ДНК- полимеразы.
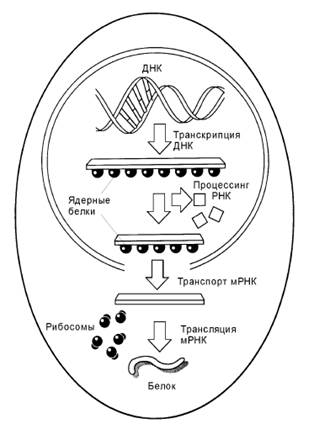
Этапы считывания генетической информации. В ходе транскрипции на ДНК-матрице синтезируется длинная молекула РНК (первичный транскрипт), содержащая последовательности экзонов и интронов. По завершении синтеза РНК-транскрипта последовательности интронов удаляются, что делает молекулу РНК значительно короче. Эта мРНК выходит из ядра в цитоплазму и соединяется с рибосомами. Молекула мРНК продвигается сквозь рибосому, и её нуклеотидная последовательность транслируется в соответствующую последовательность аминокислот создаваемой белковой цепи.
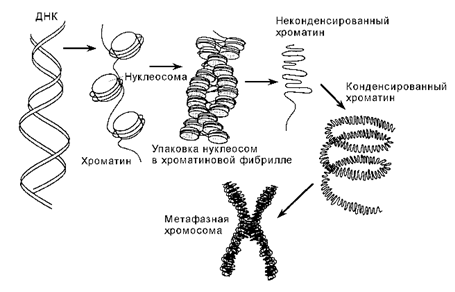
Организация хроматина в хромосоме. Хроматин состоит из структурных единиц — нуклеосом, разделённых интервалами в 200 пар оснований. Во время митоза в результате плотной упаковки нуклеосом хроматин полностью конденсируется, формируя видимые хромосомы
Нуклеосома в неконденсированном хроматине содержит по две копии гистонов H2A, H2B, H3 и H4. Двойная спираль ДНК лежит на поверхности октамера гистонов и накручена на него. В конденсированном хроматине дополнительно присутствует гистон H1, соединяющий нуклеосомы
Гранулярная эндоплазматическая сеть — система плоских мембранных цистерн с рибосомами на наружной поверхности
Cинтез белков
• для плазматической мембраны
• лизосом
• на экспорт, т.е. предназначенных для экзоцитоза
Механизм поступления белков внутрь цистерн эндоплазматической сети объясняет сигнальная гипотеза
Гладкая эндоплазматическая сеть — система анастомозирующих мембранных каналов, пузырьков и трубочек.
Не содержит рибофоринов и по этой причине не связана с рибосомами.
Функции:
• синтез стероидных гормонов
• депо Cа2+
• детоксикация
Механизм высвобождения Ca2+ из цистерн гладкой эндоплазматической сети.
А) Рецептор рианодина в мембране цистерны связан с Ca2+- каналом. Работу канала контролируют рецепторы дигидропиридина плазмолеммы, в ответ на деполяризацию активирующие рецепторы рианодина, что и приводит к высвобождению Ca2+
Б) Рецептор рианодина в мембране цистерны непосредственно не связан с белками плазмолеммы, но активируется Ca2+ при незначительном повышении концентрации этого катиона в цитозоле за счёт поступления в клетку через потенциалозависимые Ca2+ каналы
В) Ca2+ освобождается из цистерн при активации рецепторов инозитолтрифосфата. Этот второй посредник образуется при участии фосфолипазы C в ответ на взаимодействие агониста со своим рецептором в плазмолемме.
Митохондрии
• преобразователиэнергиидлявнутриклеточных реакций
• занимают значительную часть цитоплазмы клеток
• сосредоточенывместахвысокогопотребления АТФ (например, в эпителии канальцев почки они располагаются вблизи плазматической мембраны [обеспечение реабсорбции], а в нейронах — в синапсах [обеспечение электрогенеза и секреции])
Митохондрии
Генез. Предположительно произошли от аэробных симбионтов, проникших в анаэробную эукариотическую клетку путём эндоцитоза и начавших участвовать в её окислительных процессах. В связи с этим митохондрии имеют собственный геном (кольцевая ДНК), мРНК, тРНК, рРНК, но большинство белков митохондрий кодирует ядерная ДНК. Органелла функционирует в среднем 10 суток, обновление митохондрий происходит путём их деления.
Морфология. Чаще имеют форму цилиндра диаметром 0,2–1 мкм и длиной до 7 мкм. У митохондрий две мембраны — наружная и внутренняя; последняя образует кристы.
Между наружной и внутренней мембранами находится межмембранное пространство. Внемембранный объём митохондрии — матрикс.
(1) Наружная мембрана проницаема для многих мелких молекул.
(2) Межмембранное пространство. Здесь накапливаются ионы H+, выкачиваемые из матрикса, что создаёт протонный градиент концентрации по обе стороны внутренней мембраны.
(3) Внутренняя мембрана избирательно проницаема; содержит транспортные системы для переноса веществ в обоих направлениях и комплексы цепи переноса электронов, связанные с ферментами окислительного фосфорилирования, а также СДГ.
(4) Матрикс. В нём присутствуют все ферменты цикла Кребса (кроме СДГ), ферменты †b-окисления жирных кислот и некоторые ферменты других систем. В матриксе находятся гранулы с Mg и Ca.
Митохондрии: функция
(1) Окисление в цикле Кребса. В отличие от анаэробного гликолиза, в ходе которого из одной молекулы глюкозы образуется две молекулы пирувата, цикл Кребса требует присутствия O2. Гликолиз протекает в цитозоле, и образующийся пируват поступает с помощью переносчика пирувата в митохондрии в обмен на OH. Матрикс митохондрий содержит ферменты, окисляющие пируват и жирные кислоты до ацетил-КoA, и ферменты, окисляющие ацетил-КoA до CO2. Конечные продукты цикла трикарбоновых кислот (CO2, выходящий из клетки, и НАДН) — источник электронов, переносимых дыхательной цепью.
(2) Транспорт электронов. Электроны перемещаются по дыхательной цепи, локализованной во внутренней мембране и содержащей четыре крупных ферментных комплекса (преимущественно цитохромы) цепочки транспорта электронов.
(3) Фосфорилирование АДФ. Кристы митохондрий содержат АТФ-синтетазу, сопрягающую окисление в цикле Кребса и фосфорилирование АДФ до АТФ. АТФ синтезируется при обратном токе протонов в матрикс через канал в АТФ-синтезирующем комплексе.
(4) Сопряжение окисления и фосфорилирования. В результате сопряжения этих процессов энергия, освобождаемая при окислении субстратов, хранится в макроэргических связях АТФ. Освобождение энергии, запасённой в АТФ, в дальнейшем обеспечивает выполнение многочисленных функцийклеток(например,мышечноесокращение, подвижность жгутика сперматозоида, выкачивание H из париетальных клеток в железах желудка для поддержания кислой среды). Эффективность окислительного фосфорилирования в митохондриях выше эффективности гликолиза в цитозоле. Из одной молекулы глюкозы в первом случае образуется 38 молекул АТФ, а во втором — только 2.
(5) Теплопродукция. Естественный механизм разобщения окислительного фосфорилирования функционирует в клетках бурого жира. В этих клетках митохондрии имеют атипичную структуру (уменьшен их объём, увеличена плотность матрикса, расширены межмембранные пространства) — конденсированные митохондрии. Такие митохондрии могут усиленно захватывать воду и набухать в ответ на тироксин, увеличение концентрации Ca в цитозоле, при этом усиливается разобщение окислительного фосфорилирования, и происходит выделение тепла. Эти процессы обеспечивает специальный разобщающий белок. Норадреналин из симпатического отдела вегетативной нервной системы усиливает экспрессию разобщающего белка и стимулирует теплопродукцию.
(6) Контроль внутриклеточной концентрации Ca2+ — одна из важных функций митохондрий.
(7) Митохондриальный путь запуска апоптоза
Комплекс Гольджи
• Белки и липиды поступают в комплекс Гольджи с цис-стороны
• Транспортные пузырьки переносят эти молекулы последовательно из одной цистерны в другую
Готовый продукт выходит из комплекса на транс-стороне, находясь в различных пузырьках:
• часть из них содержит молекулы, предназначенные для внутриклеточного пищеварения, и сливается с лизосомами.
• другая часть пузырьков участвует в экзоцитозе.
• третья группа пузырьков содержит белки для плазмолеммы
Функции гольджи
Модификация секреторного продукта. Ферменты комплекса Гольджи гликозилируют белки и липиды; образующиеся здесь гликопротеины, протеогликаны, гликолипиды и сульфатированные гликозаминогликаны предназначены для последующей секреции
Концентрирование секреторных продуктов происходит в конденсирующих вакуолях, расположенных на транс-стороне
Упаковка секреторного продукта, образование участвующих в экзоцитозе секреторных гранул.
Аксонема
Состоит из 9 периферических пар микротрубочек и двух расположенных центрально одиночных микротрубочек. В каждой периферической паре различают субфибриллу А, содержащую 10–11 тубулиновых протофиламентов, и субфибриллу В, содержащую 13 протофиламентов. С субфибриллой А связаны наружные и внутренние ручки. В их состав входит белок динеин, обладающий АТФазной активностью. Аксонема формируется путём самосборки белковых СЕ. Матрицей для сборки служит центриоль или базальное тельце. Аксонема — основной структурный элемент реснички и жгутика.
Ресничка — вырост клетки длиной 5–10 мкм и шириной 0,2 мкм, содержащий аксонему. Реснички присутствуют в эпителиальных клетках воздухопроводящих и половых путей, перемещают слизь с инородными частицами и остатками отмерших клеток и создают ток жидкости около клеточной поверхности.
Жгутик не встречается в количестве более двух на клетку. В сперматозоиде человека имеет длину 50–55 мкм и толщину 0,2–0,5 мкм, содержит аксонему.
• Динеин. Крупный белок, содержащий 2–3 глобулярные головки, соединённые с гибкой фибриллярной частью молекулы. Основание фибриллярной части вплетено в микротрубочку (A-субфибрилла). Глобулярная головка обладает АТФазной активностью. При расщеплении АТФ она скользит по поверхности микротрубочки (B- субфибрилла) соседней пары (рис. 2-20) по направлению к её (–)-концу. Этот механизм аналогичен скольжению элементов актомиозинового хемомеханического преобразователя в мышце.
• Нарушения организации аксонемы. Дефекты ресничек и жгутиков проявляются отсутствием в аксонеме динеиновых ручек, центральной капсулы или центральных микротрубочек. Эти дефекты проявляются при синдроме неподвижных ресничек, возможно развитие рецидивирующего хронического бронхита и синусита. Более половины больных с подобным синдромом имеет situs viscerus inversus — транспозицию внутренних органов (сердце справа, печень слева и т.д.), что в совокупности описывает синдром Картагенера.
Ресничка — вырост клетки длиной 5–10 мкм и шириной 0,2 мкм, содержащий аксонему. Реснички присутствуют в эпителиальных клетках воздухопроводящих и половых путей, перемещают слизь с инородными частицами и остатками отмерших клеток и создают ток жидкости около клеточной поверхности.
Жгутик. Как правило, не встречается в количестве более двух на клетку. В сперматозоиде человека имеет длину 50–55 мкм и толщину 0,2–0,5 мкм, содержит аксонему.
Окаймлённые пузырьки
Окружены оболочкой, прилежащей к наружной поверхности мембраны, участвуют во внутриклеточной сортировке белков, образуют две разновидности — окружённые клатрином пузырьки и не содержащие клатрин пузырьки.
Окружённые клатрином пузырьки образуются в ходе опосредуемого рецепторами эндоцитоза, содержат клатриновую оболочку.
Клатриновая оболочка снаружи окружает мембрану пузырька. Главный её компонент — белок клатрин. Три его полипептидные цепи вместе с тремя цепями меньшего по размеру полипептида формируют структурный элемент оболочки — трёхвалентный белковый комплекс (трискелион); 36 подобных комплексов образует многоугольный объёмный каркас, окружающий пузырёк.
Транспортируют белки, поступающие в клетку путём опосредуемого рецепторами эндоцитоза, а также белки из транс-стороны комплекса Гольджи.
Лизосомы
· Округлые пузырьки, окружённые мембраной; их размеры и электронная плотность значительно варьируют.
· Содержат более 50 ферментов: рибонуклеазы, дезоксирибонуклеазы, катепсины, сульфатазы, глюкуронидазы, фосфолипазы, гликозидазы, липазы и другие. Лизосомные ферменты наиболее активны в кислой среде (pH–5,0), для поддержания которой в мембрану лизосомы встроен протонный насос (H,K- ATФаза).
· Участвуют во внутриклеточном пищеварении.
· При нарушении функции лизосом развиваются т.н. лизосомные болезни накопления.
Цитоскелет
Трёхмерная цитоплазматическая сеть волокнистых и трубчатых структур различного типа.
К цитоскелету относят:
· микротрубочки
· промежуточные филаменты
· микрофиламенты
· микротрабекулы
Цитоскелет придаёт клетке определённую форму и выполняет множество других функций (например, подвижность клетки, внутриклеточный транспорт)
Микротрубочки
• Полярность. Микротрубочка постоянно растет с одного конца (полимеризация) и деполимеризуется с другого конца. В микротрубочке различают (+)-конец, где присоединяются новые СЕ тубулиновых протофиламентов, и (–)-конец, где СЕ тубулина отделяются от нити. Ряд агентов блокирует сборку или деполимеризацию микротрубочек. • Ассоциированные белки.Микротрубочки ассоциированы с рядом белков, имеющих общее наименование MAP. MAP-белки стабилизируют микротрубочки и связывают их с другими элементами цитоскелета и органеллами.
• Кинезин—компоненттубулин-кинезинового хемомеханического преобразователя.
Цитостатики блокируют митоз и внутриклеточный транспорт. Это их свойство широко используют для блокады пролиферации клеток (преимущественно в онкологии).
(а) Колхицин связывается с субъединицами тубулина и препятствует их присоединению к (+)-концу микротрубочек.
(б) Алкалоиды Vinca rosea. Винбластин и его аналоги имеют тот же эффект, что и колхицин.
(в) Таксол. В отличие от алкалоидов Vinca rosea и колхицина, подавляющих сборку микротрубочек, таксол стимулирует образование стабильных микротрубочек. Это приводит к реорганизации цитоскелета с тем же конечным эффектом, что и в случае алкалоидов Vinca rosea и колхицина: происходит торможение пролиферации клеток. Таксол действует в поздней G2- и M-фазах клеточного цикла. Этот мощный ингибитор деления эукариотических клеток используют в качестве антибластомного препарата.
Функции микротрубочек
• Обеспечение расхождения хромосом при делении клеток
• Поддержание формы клетки
• Участие в транспорте макромолекул и
органелл
• Обеспечение подвижности жгутиков, ресничек
Промежуточные нити
· имеют диаметр 8–11 нм
· создают внутриклеточный каркас
· обеспечивают упругость клетки
· поддерживают упорядоченность
расположения компонентов цитоплазмы
· состоят из белков, специфичных для
определённых клеточных типов
Микрофиламенты
Имеют диаметр 6 нм.
Две переплетённые нити F-актина, составленные из G- актина.
Образуют скопления по периферии клетки и связаны с плазмолеммой посредством промежуточных белков (a- актинин, винкулин, талин).
Полярность. Как и микротрубочки, микрофиламенты полярны; присоединение (полимеризация) СЕ G-актина происходит на (+)-конце.
Токсины, связывающиеся с актином и блокирующие его полимеризацию, нарушая подвижность клеток, фагоцитоз и цитокинез.
· Цитохалазины различных плесневых грибов.
· Фаллоидин — циклический пептид бледной поганки (Amanita phalloides).
Функции Микрофиламентов:
• Изменение консистенции цитозоля, переход золя в гель и обратно (например, для изменения вязкости примембранной цитоплазмы и образования псевдоподий, активно перемещающихся в ткани нейтрофилов в ответ на хемоаттрактанты).
• Эндоцитозиэкзоцитоз.
• Подвижностьнемышечныхклетоксвязанас изменением формы клеточной поверхности вследствие регулируемой полимеризации актина.
• Стабилизациялокальныхвыпячиваний плазматической мембраны связана с пучками поперечно сшитых актиновых филаментов (например, в микроворсинках эпителиальных клеток кишки, где пучок параллельных микрофиламентов образует их сердцевину)
Включения Микрофиламенты:
Скопления в цитозоле различных гранул и капель (например, метаболически активный материал — гликоген и липиды, а также липофусцин).
Гликоген. Скопления гранул размером 20–30 нм, не связанных с мембраной, но часто расположенных вблизи гладкой эндоплазматической сети; имеет значение как энергетический резерв для образования глюкозы.
Липиды. Капли различных размеров и плотности, не связанные с мембранами; содержат триглицериды как источник энергии и холестерин, используемый для синтеза стероидов.
Липофусцин — продукт лизосомного переваривания, не подвергающийся дальнейшему внутриклеточному расщеплению (мембранные пузырьки различных размеров с электроноплотным содержимым, часто содержат липиды). Другое название — пигмент старения.
Цитозоль
Цитозоль — жидкая часть цитоплазмы, составляет около половины объёма клетки. Здесь синтезируются белки, часть которых собирается на полисомах и остаётся в цитозоле (например, Hb, протеинкиназы). В синтезе цитозольных белков участвуют мРНК, не имеющие сигнальных кодонов. В цитозоле присутствуют нелизосомные протеазы, переваривающие короткоживущие белки.
Клеточный цикл
Фазы
G1 — обычно самая продолжительная фаза цикла, следует за телофазой митоза. В эту фазу клетка синтезирует РНК и белки. Продолжительность фазы — от нескольких часов до нескольких дней. У быстро делящихся клеток (эмбриональные и неопластические) эта фаза непродолжительна.
G0. Клетки могут выйти из цикла и находиться в фазе G0. Они начинают дифференцироваться, достигая состояния терминальной (окончательной) дифференцировки (например, нейроны).
S. В фазу S в клетке продолжается синтез белка, происходит репликация ДНК, разделяются центриоли. В большинстве клеток фаза S длится 8–12 часов.
G2. В фазу G2 продолжается синтез РНК и белка (например, синтез тубулина для микротрубочек митотического веретена). Центриоли достигают размеров дефинитивных органелл. В эту же фазу накапливается АТФ для энергетического обеспечения последующего митоза. Эта фаза длится 2–4 часа.
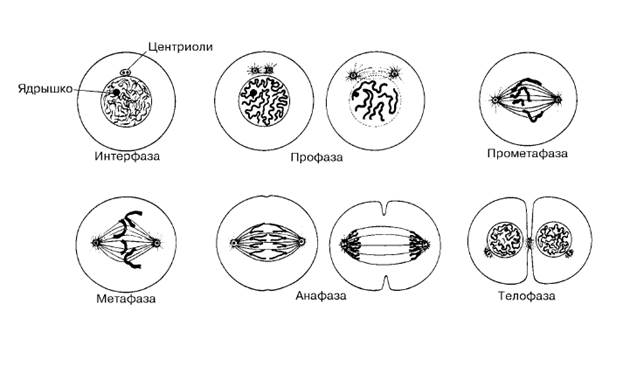
Митоз
В ходе митоза делятся ядро (кариокинез) и цитоплазма (цитокинез).
Фазы митоза:
Профаза
· Конденсация хромосом. Каждая хромосома состоит из двух сестринских хроматид, соединённых центромерой.
· Исчезновение ядрышка.
· Образование митотического веретена. Прометафаза
· Ядерная оболочка распадается на мелкие фрагменты.
· В области центромер появляются кинетохоры, функционирующие как центры организации кинетохорных микротрубочек. Отхождение кинетохор от каждой хромосомы в обе стороны и их взаимодействие с полюсными микротрубочками митотического веретена — причина перемещения хромосом.
Метофаза
• Хромосомы располагаются в области экватора веретена.
• Образуется метафазная пластинка, в которой каждая хромосома удерживается парой кинетохоров и связанными с ними кинетохорными микротрубочками, направленными к противоположным полюсам митотического веретена.
Анафаза — расхождение дочерних хромосом к полюсам митотического веретена со скоростью 1 мкм/мин.
Телофаза
· Хроматидыподходяткполюсам, кинетохорные микротрубочки исчезают, а полюсные продолжают удлиняться.
· Образование ядерной оболочки.
· Появление ядрышка.
Цитокинез — разделение цитоплазмы на две обособляющиеся части.
Основные компартменты клетки:
Клетка — главный гистологический элемент
Три основных компартмента:
· плазматическая мембрана
· ядро
· цитоплазма
Органеллы, включения
Биологические мембраны входят в состав каждого клеточного компартмента и многих органелл.
Компартмент - отдельная, функционально значимая часть клетки или обособленная субклеточная структура, характеризующаяся специфическими морфологическими или биохимическими свойствами и отделённая от остальной клетки по крайней мере одной избирательно проницаемой мембраной.
Химический состав (плазмолемма, клеточная мембрана):
1. Липиды
2. Белки (50%)
3. Углеводы (2–10%)
Функции:
· избирательная проницаемость
· межклеточные взаимодействия
· эндоцитоз
· экзоцитоз
Избирательная проницаемость - поддерживает клеточный гомеостаз, оптимальное содержание в клетке ионов, воды, ферментов и субстратов.
Пути реализации:
• пассивный транспорт
• облегчённая диффузия
• активный транспорт
Ионные каналы (натриевые, калиевые, Ca, хлорные каналы, водные каналы) (аквапорины)
Активный транспорт — энергозависимый трансмембранный перенос против электрохимического градиента, происходящий при участии АТФаз
Натрий, калиевая АТФаза (Na,K-АТФаза) Протонная и калиевая АТФаза (H,K-АТФаза) Са-транспортирующие АТФазы (Ca-АТФаза)
Межклеточные взаимодействия
Плазматическая мембрана — место приложения раздражителей
Из внешней среды:
• физических(например, кванты света в фоторецепторах)
• химических(например, вкусовые и обонятельные молекулы, рН)
• механических(например, давление или растяжение в механорецепторах)
Из внутренней среды организма:
сигналов информационного характера (например, гормоны, нейромедиаторы)
Эндоцитоз — поглощение (интернализация) клеткой веществ, частиц и микроорганизмов.
Варианты эндоцитоза:
· Пиноцитоз — процесс поглощения жидкости и растворённых веществ с образованием небольших пузырьков. Пиноцитозные пузырьки формируются в специализированных областях плазматической мембраны — окаймлённых ямках
· Фагоцитоз — поглощение крупных частиц (например, микроорганизмов или остатков клеток).
· Опосредуемый рецепторами эндоцитоз характеризуется поглощением из внеклеточной жидкости конкретных макромолекул.
·
Опосредуемый рецепторами эндоцитоз. Многие внеклеточные макромолекулы (трансферрин, ЛНП, вирусные частицы и др.) связываются со своими рецепторами в плазмолемме. Образуются клатриновые окаймлённые ямки, а затем — окаймлённые пузырьки, содержащие комплекс лиганд-рецептор. Окаймлённые пузырьки после освобождения от клатрина — эндосома. Внутри эндосом лиганд отщепляется от рецептора.
Экзоцитоз — процесс, при котором внутриклеточные секреторные пузырьки (например, синаптические) и секреторные гранулы сливаются с плазмолеммой, а их содержимое освобождается из клетки — секреция .
Спонтанная и регулируемая секреция. Одна часть пузырьков постоянно сливается с клеточной мембраной (спонтанная секреция), в то время как другая часть скапливается под ней, и процесс слияния происходит только под действием сигнала, чаще всего вследствие увеличения концентрации Са2+ в цитозоле (регулируемый экзоцитоз).
1. Спонтанная секреция обеспечивает встраивание в плазмолемму вновь синтезированных белков и рецепторов, интернализованных при опосредованном рецепторами эндоцитозе.
2. Регулируемый экзоцитоз. В этом процессе участвуют секреторные гранулы, а также специализированные эндосомы (например, синаптические пузырьки).
Ядро
23 пары хромосом (диплоидный набор) ядра соматической клетки содержат ядерный геном (примерно 20-30.000 генов).
Реализация генетической информации (транскрипция ® процессинг ® трансляция ® посттрансляционная модификация) и другие функции ядра происходят при участии ДНК и разных видов РНК.
Структурные компоненты ядра: хроматин, ядерная оболочка, ядрышко, нуклеоплазма.
ДНК состоит из двух полинуклеотидных цепей, образованных нуклеотидами, соединёнными фосфодиэфирными связями (1). Ковалентные фосфодиэфирные связи соединяют 5’- атом углерода одного нуклеотида с 3’- атомом углерода следующего нуклеотида. Антипараллельные цепи комплементарно спарены (2): аденин (A) с тимином (T), гуанин (G) c цитозином (C). Клетки перед каждым делением воспроизводят (реплицируют) ДНК (3): дочерние молекулы ДНК воспроизводятся при помощи ДНК- полимеразы.
Этапы считывания генетической информации. В ходе транскрипции на ДНК-матрице синтезируется длинная молекула РНК (первичный транскрипт), содержащая последовательности экзонов и интронов. По завершении синтеза РНК-транскрипта последовательности интронов удаляются, что делает молекулу РНК значительно короче. Эта мРНК выходит из ядра в цитоплазму и соединяется с рибосомами. Молекула мРНК продвигается сквозь рибосому, и её нуклеотидная последовательность транслируется в соответствующую последовательность аминокислот создаваемой белковой цепи.
Организация хроматина в хромосоме. Хроматин состоит из структурных единиц — нуклеосом, разделённых интервалами в 200 пар оснований. Во время митоза в результате плотной упаковки нуклеосом хроматин полностью конденсируется, формируя видимые хромосомы
Нуклеосома в неконденсированном хроматине содержит по две копии гистонов H2A, H2B, H3 и H4. Двойная спираль ДНК лежит на поверхности октамера гистонов и накручена на него. В конденсированном хроматине дополнительно присутствует гистон H1, соединяющий нуклеосомы
Гранулярная эндоплазматическая сеть — система плоских мембранных цистерн с рибосомами на наружной поверхности
Cинтез белков
• для плазматической мембраны
• лизосом
• на экспорт, т.е. предназначенных для экзоцитоза
Механизм поступления белков внутрь цистерн эндоплазматической сети объясняет сигнальная гипотеза
Гладкая эндоплазматическая сеть — система анастомозирующих мембранных каналов, пузырьков и трубочек.
Не содержит рибофоринов и по этой причине не связана с рибосомами.
Функции:
• синтез стероидных гормонов
• депо Cа2+
• детоксикация
Механизм высвобождения Ca2+ из цистерн гладкой эндоплазматической сети.
А) Рецептор рианодина в мембране цистерны связан с Ca2+- каналом. Работу канала контролируют рецепторы дигидропиридина плазмолеммы, в ответ на деполяризацию активирующие рецепторы рианодина, что и приводит к высвобождению Ca2+
Б) Рецептор рианодина в мембране цистерны непосредственно не связан с белками плазмолеммы, но активируется Ca2+ при незначительном повышении концентрации этого катиона в цитозоле за счёт поступления в клетку через потенциалозависимые Ca2+ каналы
В) Ca2+ освобождается из цистерн при активации рецепторов инозитолтрифосфата. Этот второй посредник образуется при участии фосфолипазы C в ответ на взаимодействие агониста со своим рецептором в плазмолемме.
Митохондрии
• преобразователиэнергиидлявнутриклеточных реакций
• занимают значительную часть цитоплазмы клеток
• сосредоточенывместахвысокогопотребления АТФ (например, в эпителии канальцев почки они располагаются вблизи плазматической мембраны [обеспечение реабсорбции], а в нейронах — в синапсах [обеспечение электрогенеза и секреции])
Митохондрии
Генез. Предположительно произошли от аэробных симбионтов, проникших в анаэробную эукариотическую клетку путём эндоцитоза и начавших участвовать в её окислительных процессах. В связи с этим митохондрии имеют собственный геном (кольцевая ДНК), мРНК, тРНК, рРНК, но большинство белков митохондрий кодирует ядерная ДНК. Органелла функционирует в среднем 10 суток, обновление митохондрий происходит путём их деления.
Морфология. Чаще имеют форму цилиндра диаметром 0,2–1 мкм и длиной до 7 мкм. У митохондрий две мембраны — наружная и внутренняя; последняя образует кристы.
Между наружной и внутренней мембранами находится межмембранное пространство. Внемембранный объём митохондрии — матрикс.
(1) Наружная мембрана проницаема для многих мелких молекул.
(2) Межмембранное пространство. Здесь накапливаются ионы H+, выкачиваемые из матрикса, что создаёт протонный градиент концентрации по обе стороны внутренней мембраны.
(3) Внутренняя мембрана избирательно проницаема; содержит транспортные системы для переноса веществ в обоих направлениях и комплексы цепи переноса электронов, связанные с ферментами окислительного фосфорилирования, а также СДГ.
(4) Матрикс. В нём присутствуют все ферменты цикла Кребса (кроме СДГ), ферменты †b-окисления жирных кислот и некоторые ферменты других систем. В матриксе находятся гранулы с Mg и Ca.
Митохондрии: функция
(1) Окисление в цикле Кребса. В отличие от анаэробного гликолиза, в ходе которого из одной молекулы глюкозы образуется две молекулы пирувата, цикл Кребса требует присутствия O2. Гликолиз протекает в цитозоле, и образующийся пируват поступает с помощью переносчика пирувата в митохондрии в обмен на OH. Матрикс митохондрий содержит ферменты, окисляющие пируват и жирные кислоты до ацетил-КoA, и ферменты, окисляющие ацетил-КoA до CO2. Конечные продукты цикла трикарбоновых кислот (CO2, выходящий из клетки, и НАДН) — источник электронов, переносимых дыхательной цепью.
(2) Транспорт электронов. Электроны перемещаются по дыхательной цепи, локализованной во внутренней мембране и содержащей четыре крупных ферментных комплекса (преимущественно цитохромы) цепочки транспорта электронов.
(3) Фосфорилирование АДФ. Кристы митохондрий содержат АТФ-синтетазу, сопрягающую окисление в цикле Кребса и фосфорилирование АДФ до АТФ. АТФ синтезируется при обратном токе протонов в матрикс через канал в АТФ-синтезирующем комплексе.
(4) Сопряжение окисления и фосфорилирования. В результате сопряжения этих процессов энергия, освобождаемая при окислении субстратов, хранится в макроэргических связях АТФ. Освобождение энергии, запасённой в АТФ, в дальнейшем обеспечивает выполнение многочисленных функцийклеток(например,мышечноесокращение, подвижность жгутика сперматозоида, выкачивание H из париетальных клеток в железах желудка для поддержания кислой среды). Эффективность окислительного фосфорилирования в митохондриях выше эффективности гликолиза в цитозоле. Из одной молекулы глюкозы в первом случае образуется 38 молекул АТФ, а во втором — только 2.
(5) Теплопродукция. Естественный механизм разобщения окислительного фосфорилирования функционирует в клетках бурого жира. В этих клетках митохондрии имеют атипичную структуру (уменьшен их объём, увеличена плотность матрикса, расширены межмембранные пространства) — конденсированные митохондрии. Такие митохондрии могут усиленно захватывать воду и набухать в ответ на тироксин, увеличение концентрации Ca в цитозоле, при этом усиливается разобщение окислительного фосфорилирования, и происходит выделение тепла. Эти процессы обеспечивает специальный разобщающий белок. Норадреналин из симпатического отдела вегетативной нервной системы усиливает экспрессию разобщающего белка и стимулирует теплопродукцию.
(6) Контроль внутриклеточной концентрации Ca2+ — одна из важных функций митохондрий.
(7) Митохондриальный путь запуска апоптоза
Комплекс Гольджи
• Белки и липиды поступают в комплекс Гольджи с цис-стороны
• Транспортные пузырьки переносят эти молекулы последовательно из одной цистерны в другую
Готовый продукт выходит из комплекса на транс-стороне, находясь в различных пузырьках:
• часть из них содержит молекулы, предназначенные для внутриклеточного пищеварения, и сливается с лизосомами.
• другая часть пузырьков участвует в экзоцитозе.
• третья группа пузырьков содержит белки для плазмолеммы
Дата: 2019-02-18, просмотров: 1113.