Возбудитель СПИДа — вирус иммунодифицита человека (ВИЧ) — относится к ретровирусам[40]. Вирион имеет сферическую форму, диаметром 100-150 нм (рис. 322). Наружная оболочка вируса состоит из мембраны, образованной из клеточной мембраны клетки-хозяина.
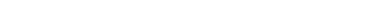
Поэтому на ее поверхности и внутри нее сохраняется множество клеточных белков. В мембрану встроены рецепторные образования, по виду напоминающие грибы. Под наружной оболочкой располагается сердцевина вируса, которая имеет форму усеченного конуса и образована особым белком. Промежуток между наружной вирусной мембраной и сердцевиной вируса заполнен тяжами вироскелета, благодаря которому сохраняется форма вируса, а сердцевина удерживается в определенном положении. Внутри сердцевины располагаются две молекулы вирусной РНК, связанные с низкомолекулярными белками основного характера. Каждая моле кула РНК содержит 9 генов ВИЧ. Три из них являются структурными, три — регуляторными и три — дополнительными. Эти гены содержат информацию, необходимую для продукции белков, которые управляют способностью вируса инфицировать клетку, реплицироваться и вызывать заболевание. Кроме того, сердцевина содержит фермент обратную транскриптазу, осуществляющую синтез вирусной ДНК с молекулы вирусной РНК.
|
Пути распространения ВИЧ инфекции |
Источником заражения служит человек — носитель вируса иммунодефицита. Это может быть больной с различными проявлениями болезни, или человек, не имеющий признаков заболевания (бессимптомный вирусоноситель).
СПИД передается только от человека к человеку:
© половым путем;
© через кровь и ткани, содержащие вирус иммунодефицита;
© от матери к плоду и новорожденному.
Глава 40. Основы генетики и селекции
Введение в генетику
Предмет генетики
Генетика — относительно молодая наука. Официальной датой ее рождения считается 1900г., когда Г. де Фриз в Голландии, К.Корренс в Германии и Э.Чермак в Австрии независимо друг от друга "переоткрыли" законы наследования признаков, установленные Г. Менделем еще в 1865 году.
Генетика изучает два фундаментальных свойства живых организмов: наследственность и изменчивость.
Под наследственностью понимают свойство организмов обеспечивать материальную и функциональную преемственность между поколениями. Благодаря наследственности, каждый вид животных и растений в ряде сменяющих друг друга поколений сохраняет не только характерные для него признаки, но и особенности развития.
Материальной основой наследственности, связывающей поколения, являются клетки — гаметы (при половом размножении) и соматические (при бесполом). Но клетки несут в себе не признаки и свойства будущих организмов, а лишь задатки, дающие возможность развития этих признаков и свойств. Этими задатками являются гены. Эукариотическим геном называют совокупность сегментов молекулы ДНК, которая дает начало или молекуле РНК, или полипептиду. Совокупность всех генов организма называют генотипом.
Наличие задатка еще не означает обязательного появления признака, поскольку развитие любого признака зависит как от присутствия других генов, так и от условий среды. То есть, формирование признаков происходит в ходе индивидуального развития особей. Поэтому каждая отдельно взятая особь уникальна, так как обладает набором признаков, характерных только для нее.
Совокупность всех признаков организма называют фенотипом. Сюда относятся не только видимые признаки (цвет глаз, волос и т.д.), но и биохимические (структура белков, активность ферментов и т.д.), гистологические (форма и размеры клеток, строение тканей и органов), анатомические (строение тела и взаимное расположение органов). То есть, признаком может быть названа любая особенность строения, любое свойство организма.
Появление в рамках одного вида признаков, отличающих особей друг от друга, является следствием наличия у особей свойства изменчивости. Под изменчивостью понимают свойство организмов приобретать новые признаки под воздействием различных факторов. Изменчивость заключается в изменении наследственных задатков, то есть генов. Изучением причин и форм изменчивости также занимается генетика.
Изменчивость противоположна наследственности. Если наследственность стремится закрепить признаки и свойства организмов, то изменчивость обеспечивает появление новых признаков и свойств. Вмести с тем, наследственность и изменчивость тесно взаимосвязаны. Благодаря изменчивости организмы приспосабливаются к изменяющимся условиям окружающей среды, а благодаря наследственности эти изменения закрепляются.
Таким образом, генетика — это наука о закономерностях наследственности и изменчивости.
|
Методы генетики |
Как любая наука, генетика имеет свои методы исследования. Основным является г ибридологический метод — система скрещиваний, позволяющая проследить закономерности наследования и изменения признаков в ряду поколений. Метод разработан Г. Менделем. От обычных скрещиваний с последующим наблюдением за потомством этот метод отличается следующими особенностями:
¨ целенаправленный подбор родителей, различающихся по одной, двум, трем и т.д. парам контрастных (альтернативных) стабильных признаков;
¨ учет наследования признаков отдельно по каждой такой паре в каждом поколении;
¨ строгий количественный учет наследования признаков у гибридов ряда последовательных поколений;
¨ индивидуальная оценка потомства от каждого родителя в ряду поколений.
Другие методы будут подробно рассмотрены при изучении следующих тем. К ним относятся: генеалогический — составление и анализ родословных; цитогенетический — изучение хромосом при помощи микроскопа; близнецовый — изучение генетических закономерностей на близнецах; популяционно-статистический метод — изучение генетической структуры популяций.
|
Генетическая символика |
Для записи результатов скрещиваний в генетике используются специальная символика, предложенная Г.Менделем:
© Р — родители;
© F — потомство, число внизу или сразу после буквы указывает на порядковый номер поколения (F1 — гибриды первого поколения — прямые потомки родителей, F2 — гибриды второго поколения — возникают в результате скрещивания между собой гибридов F1);
© х — значок скрещивания;
© ♂ — мужская особь;
© ♀ — женская особь
© A, a, B, b, C, c — буквами латинского алфавита обозначаются отдельно взятые наследственные признаки.
Законы Менделя
|
 Грегор Иоганн Мендель
(1822 — 1884)
Грегор Иоганн Мендель
(1822 — 1884)
© относительно просто выращивается и имеет короткий период развития, что позволяет достаточно быстро получить потомство от скрещивания, причем за год можно получить несколько поколений;
© имеет многочисленное потомство, что удобно для проведения статистического анализа;
© имеет большое количество хорошо заметных альтернативных признаков:
¨ окраска венчика — белая или красная;
¨ окраска семядолей — зеленая или желтая;
¨ форма семени — морщинистая или гладкая;
¨ окраска боба — желтая или зеленая;
¨ форма боба — округлая или с перетяжками;
¨ расположение цветков или плодов — по всей длине стебля или у его верхушки;
¨ высота стебля — длинный или короткий;
© является самоопылителем, в результате чего имеет большое количество чистых линий, устойчиво сохраняющих свои признаки из поколения в поколение;
© строение венчика цветка позволяет защитить цветок от опыления посторонней пыльцой.
Опыты Менделя были тщательно продуманы. Если его предшественники пытались изучить закономерности наследования сразу многих признаков, то Мендель шел от простого к сложному. Свои исследования он начал с изучения закономерностей наследования всего лишь одной пары альтернативных признаков.
Моногибридным называют скрещивание двух организмов, отличающихся друг от друга по одной паре альтернативных (взаимоисключающих) признаков. Таким образом, при таком скрещивании прослеживаются закономерности наследования только двух вариантов признака (например, белая и красная окраска венчика), а все остальные признаки организма во внимание не принимаются.
|
Первый закон Менделя |
Классическим примером моногибридного скрещивания является скрещивание сортов гороха с желтыми и зелеными семенами (рис. 323). При скрещивании растения с желтыми и зелеными семе-
|
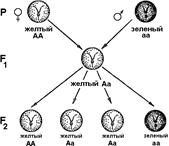 Рис. 323. Моногибридное скрещивание растений гороха с желтыми и зелеными семенами.
Рис. 323. Моногибридное скрещивание растений гороха с желтыми и зелеными семенами.
как бы исчезает. Проявляющийся у гибридов первого поколения признак Мендель назвал доминантным, а подавляемый — рецессивным. Само же явление преобладания у гибридов признака одного из родителей Г. Мендель назвал доминированием.
Позже выявленная закономерность была названа законом единообразия гибридов первого поколения, или законом доминирования. Это первый закон Менделя: при скрещивании двух организмов, относящихся к разным чистым линиям (двух гомозиготных организмов), отличающихся друг от друга по одной паре альтернативных признаков, все первое поколение гибридов ( F1) окажется единообразным и будет нести признак одного из родителей.
|
Второй закон Менделя |
Второй закон наследственности был сформулирован Менделем при изучении гибридов второго поколения. Семена гибридов первого поколения использовались Менделем для получения второго гибридного поколения. Результаты опытов Менделя приведены в таблице.
Таблица 8
Результаты расщепления по различным признакам в F2,
полученные в опытах Г.Менделя с горохом.
| Признаки | Доминантные | Рецессивные | Всего | ||
| число | % | число | % | ||
| Форма семян | 5474 | 74,74 | 1850 | 25,26 | 7324 |
| Окраска семядолей | 6022 | 75,06 | 2001 | 24,94 | 8023 |
| Окраска семенной кожуры | 705 | 75,90 | 224 | 24,10 | 929 |
| Форма боба | 882 | 74,68 | 299 | 25,32 | 1181 |
| Окраска боба | 428 | 73,79 | 152 | 26,21 | 580 |
| Расположение цветков | 651 | 75,87 | 207 | 24,13 | 858 |
| Высота стебля | 787 | 73,96 | 277 | 26,04 | 1064 |
| Всего: | 14949 | 74,90 | 5010 | 25,10 | 19959 |
Анализ данных таблицы позволяет сделать ряд выводов:
© единообразия гибридов во втором поколении не наблюдается — часть гибридов несет один (доминантный), часть — другой (рецессивный) признак из альтернативной пары;
© количество гибридов, несущих доминантный признак, приблизительно в 3 раза больше, чем гибридов, несущих рецессивный признак, причем это соотношение наблюдается и по каждой отдельно взятой паре, и по всей совокупности растений;
© рецессивный признак не исчезает, а лишь подавляется и проявляется во втором гибридном поколении;
© наследуются не сами признаки, а наследственные задатки, или факторы (в современной терминологии — гены), их определяющие.
Явление, при котором часть гибридов второго поколения несет доминантный признак, а часть — рецессивный, называют расщеплением. Причем наблюдающееся у гибридов расщепление не случайное, а подчиняется определенным количественным закономерностям.
Таким образом, на основе скрещивания гибридов первого поколения и анализа второго был сформулирован второй закон Менделя: при скрещивании гибридов первого поколения в потомстве происходит расщепление признаков в определенном числовом соотношении.
|
Гипотеза чистоты гамет |
Для объяснения явления доминирования и расщепления гибридов второго поколения Мендель предложил гипотезу чистоты гамет. Он предположил, что развитие признака определяется соответствующим ему наследственным фактором. Один наследственный фактор гибриды получают от отца, другой — от матери. У гибридов F1 проявляется лишь один из факторов — доминантный. Однако, среди гибридов F2, появляются особи с признаками исходных родительских форм. Это значит, что наследственные факторы сохраняются в неизменном виде, а в половые клетки попадает только один наследственный фактор, то есть они "чисты" (не содержат второго наследственного фактора).
Итак, гипотеза чистоты гамет гласит: гаметы "чисты", содержат только один наследственный признак из пары.
|
Аллелизм |
Наследственные задатки (гены) Мендель предложил обозначать большими буквами латинского алфавита, например, доминантный — большой — А, рецессивный — маленькой — а.
Каждый организм один задаток (ген) получает от материнского организма, а другой — от отцовского, следовательно, они являются парами. Явление парности генов называют аллелизмом, парные гены — аллельными, а каждый ген пары — аллелью. Например, желтая и зеленая окраска семян гороха являются двумя аллелями (соответственно, доминантный аллель и рецессивный аллель) одного гена.
|
Множественный аллелизм |
|
Цитологические основы Моногибридного скрещивания |
В настоящее время известно, что существуют гены, имеющие не два, а большее количество аллелей. Например, у мухи дрозофилы ген окраски глаз представлен 12 аллелями: красная, коралловая, вишневая, абрикосовая и т.д. до белой. Наличие у гена большого количества аллелей называют множественным аллелизмом. Множественный аллелизм является следствием возникновения нескольких мутаций одного и того же гена.
Поскольку в своих опытах Г. Мендель использовал растения, относящиеся к разным чистым линиям, аллельные гены этих растений одинаковы. Организмы, имеющие одинаковые аллели одного гена, называются гомозиготными. Они могут быть гомозиготными по доминантным (АА) или по рецессивным генам (аа). Организмы, имеющие разные аллели одного гена, называются гетерозиготными (Аа).
Во времена Менделя строение и развитие половых клеток еще не было изучено. Поэтому его гипотеза чистоты гамет является примером гениального предвидения, которое позже нашло научное подтверждение.
Явления доминирования и расщепления признаков, наблюдавшиеся Менделем, в настоящее время легко объясняются парностью хромосом, расхождением хромосом во время мейоза и объединением их во время оплодотворения (рис. 324).
|
 Рис. 324. Расхождение хромосом при мейозе.
Рис. 324. Расхождение хромосом при мейозе.
Генетическая запись осуществляется следующим образом:
Дано: Решение:
 Р АА х аа
Р АА х аа
Ген Признак Желт. Зелен.
 А — желтые семена; Гам.
А — желтые семена; Гам.
а — зеленые семена;
Р АА х аа F1 Аа х Аа
Желт. Зелен. Желт. Желт.
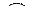
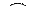
 F1 = ? Гам.
F1 = ? Гам.
F2 АА + 2Аа + аа
Желт. Желт. Зелен.
При оплодотворении гаметы сливаются, и их хромосомы объединяются в одной зиготе. Получившийся от скрещивания гибрид становится гетерозиготным, так как его клетки будут иметь генотип Аа, то есть оба аллеля одного и того же гена. У гибридного организма во время мейоза хромосомы расходятся в разные клетки и образуется два типа гамет — 50% гамет будет нести ген А, 50% — ген а. Оплодотворение — процесс случайный и равновероятный, то есть любой сперматозоид может оплодотворить любую клетку. А поскольку образовалось два типа сперматозоидов и два типа яйцеклеток, возможно возникновение четырех типов зигот.
Для удобства расчета сочетания гамет при оплодотворении английский генетик Р.Пеннет предложил проводить запись в виде решетки, которую так и назвали — решетка Пеннета. По вертикали указываются женские гаметы, по горизонтали — мужские. В клетки решетки вписываются генотипы зигот, образовавшихся при слиянии гамет.
Из приведенной схемы видно, что образуется три типа зигот. Половина из них — гетерозиготы (несут гены А и а), 1/4 — гомозиготы по доминантному признаку (несут два гена А) и 1/4 — гомозиготы по рецессивному признаку (несут два гена а). Причем желтосеменные растения одинаковы по фенотипу, но различны по генотипу: 1/3 являются гомозиготными по доминантному признаку и 2/3 — гетерозиготны.
Таким образом, учитывая цитологические основы, второй закон Менделя можно сформулировать следующим образом: при скрещивании гибридов первого поколения между собой (двух гетерозиготных особей) во втором поколении наблюдается расщепление в определенном числовом соотношении: по фенотипу 3:1, по генотипу 1:2:1.
|
Неполное доминирование |
Явление доминирования не абсолютно. Сам Мендель столкнулся с тем, что при скрещивании крупнолистного сорта гороха с мелколистным гибриды первого поколения не повторяли признак ни одного из родительских растений. Все они имели листья средней величины, то есть выражение признака у гибридов носит промежуточный характер с большим или меньшим уклонением в сторону одного из родительских признаков.
|
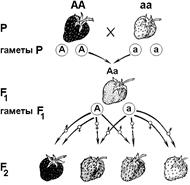 Рис. 326. Наследование окраски плода у земляники при явлении неполного доминирования.
Рис. 326. Наследование окраски плода у земляники при явлении неполного доминирования.
В качестве примера рассмотрим наследование окраски плода у земляники (рис. 326). При скрещивании гомозиготных красноплодных и белоплодных сортов земляники, все первое поколение гибридов получается розовоплодным. При скрещивании гибридов получаем
расщепление в соотношении соотношении 1 красноплодная: 2 розовоплодные: 1 белоплодная. Характерно то, что при неполном доминировании расщепление по генотипу соответствует расщеплению по фенотипу, так как гетерозиготы фенотипически отличаются от гомозигот.
Анализирующее скрещивание. Генотип гороха с зелеными семенами может быть только аа. Горох с желтыми семенами может иметь генотип АА или Аа. Для того, чтобы определить генотип особи, обладающей доминантными признаками, проводят анализирующее скрещивание — скрещивают с особью, гомозиготной по рецессивным признакам.
Если исследуемая особь гомозиготна (АА), то потомство от такого скрещивания будет иметь желтые семена и генотип Аа:
АА х аа; F1 — 100% Аа.
Если исследуемая особь гетерозиготна (Аа), то она образует два типа гамет и 50% потомства будет иметь желтые семена и генотип Аа, а 50% — зеленые семена и генотип аа: Аа х аа; F1 — 50% Аа, 50% аа.
|
Дигибридное скрещивание |
Организмы отличаются друг от друга по многим признакам. Поэтому, установив закономерности наследования одной пары признаков, Г.Мендель перешел к изучению наследования двух (и более) пар альтернативных признаков.
Дигибридным называют скрещивание двух организмов, отличающихся друг от друга по двум парам альтернативных признаков.
|
Третий закон Менделя |
Для дигибридного скрещивания Мендель брал гомозиготные растения гороха, отличающиеся по окраске семян (желтые и зеленые) и форме семян (гладкие и морщинистые). Желтая окраска (А) и гладкая форма (В) семян — доминантные признаки, зеленая окраска (а) и морщинистая форма (в) — рецессивные признаки.
Скрещивая растение с желтыми и гладкими семенами с растением с зелеными и морщинистыми семенами, Мендель получил единообразное гибридное поколение F1 с желтыми и гладкими семенами (рис. 325). От самоопыления 15 гибридов F1 было получено 556 семян, из них 315 желтых гладких, 101 желтое морщинистое, 108 зеленых гладких и 32 зеленых морщинистых.
Анализируя полученное потомство, Мендель, прежде всего, обратил внимание на то, что, наряду с сочетаниями признаков исходных сортов (желтые гладкие и зеленые морщинистые семена), при дигибридном скрещивании появляются и новые сочетания признаков (желтые морщинистые и зеленые гладкие семена). Он обратил внимание на то, что расщепление по каждому отдельно взятому признаку соответствует расщеплению при моногибридном скрещивании. Из 556 семян 423 были гладкими и 133 морщинистыми (соотношение 3:1), 416 семян имели желтую окраску, а 140 — зеленую (соотношение 3:1). Однако Менделя интересовал вопрос: зависит ли расщепление одной пары признаков (гладкие и морщинистые семена) от расщепления другой пары (желая окраска семян и зеленая) или эти пары тесно связаны между собой.
|
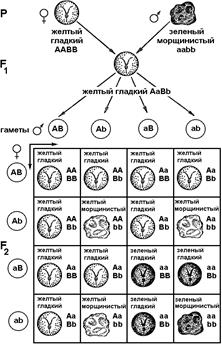 Рис. 325. Дигибридное скрещивание растений гороха с желтыми гладкими и зелеными морщинистыми семенами.
Рис. 325. Дигибридное скрещивание растений гороха с желтыми гладкими и зелеными морщинистыми семенами.
© 9/16 растений F2 обладали обоими доминантными признаками (гладкие желтые семена);
© 3/16 были желтыми (доминантный) и морщинистыми (рецессивный);
© 3/16 были зелеными (рецессивный) и гладкими (доминантный);
© 1/16 растений F2 обладали обоими рецессивными признаками (морщинистые семена зеленого цвета).
Если при моногибридном скрещивании родительские организмы отличаются по одной паре признаков (21) (желтые и зеленые семена) и дают во втором поколении два
фенотипа в соотношении 3+1, то при дигибридном они отличаются по двум парам признаков (22) и дают во втором поколении четыре фенотипа в соотношении (3+1)2. Легко посчитать, сколько фенотипов и в каком соотношении будет образовываться во втором поколении при тригибридном скрещивании: (23). — восемь фенотипов в соотношении (3+1)3.
Четыре фенотипа скрывают девять разных генотипов: 1 — ААBB; 2 — AABb; 1 — AAbb; 2 — AaBB; 4 — AaBb; 2 — Aabb; 1 — aaBB; 2 — aaBb; 1 — aabb. Если расщепление по генотипу в F2 при моногибридном поколении было 1:2:1, то есть было три разных генотипа (31), то при при дигибридном образуется 9 разных генотипов — 32, при тригибридном скрещивании образуется 33 — 27 разных генотипов.
Мендель пришел к выводу, что расщепление по одной паре признаков не связано с расщеплением по другой паре. Для семян гибридов характерны не только сочетания признаков родительских растений (желтое гладкое семя и зеленое морщиностое семя), но и возникновение новых комбинаций признаков (желтое морщинистое семя и зеленое гладкое семя).
Проведенное исследование позволило сформулировать закон независимого комбинирования генов (третий закон Менделя): при скрещивании двух гетерозиготных особей, отличающихся друг от друга по двум (и более) парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга в соотношении 3:1 и комбинируются во всех возможных сочетаниях.
Третий закон Менделя справедлив только для тех случаев, когда анализируемые гены находятся в разных парах гомологичных хромосом.
|
Цитологические основы Третьего закона Менделя |
Пусть А — ген, обусловливающий развитие желтой окраски семян, а — зеленой окраски, В — гладкая форма семени, в — морщинистая. Скрещиваются гибриды первого поколения, имеющие генотип АаВв. При образовании гамет, из каждой пары аллельных генов в гамету попадает только один, при этом в результате случайности расхождения хромосом в первом делении мейоза ген А может попасть в одну гамету с геном В или с геном в, а ген а может объединиться с геном В или с геном в. Таким образом, каждый организм образует четыре сорта гамет в одинаковом количестве (по 25 %): АВ, Aв, aB, aв. Во время оплодотворения каждый из четырех типов сперматозоидов может оплодотворить любую из четырех типов яйцеклеток. Все возможные сочетания мужских и женских гамет легко установить с помощью решетки Пеннета. При анализе результатов видно, что по фенотипу потомство делится на четыре группы: 9 желтых гладких: 3 желтых морщинистых: 3 зеленых гладких: 1 желтая морщинистая. Если проанализировать результаты расщепления по каждой паре признаков в отдельности, то получится, что отношение числа желтых семян к числу зеленых — 3:1, отношение числа гладких к числу морщинистых — 3:1. Таким образом, при дигибридном скрещивании каждая пара признаков при расщеплении в потомстве ведет себя так же, как при моногибридном скрещивании, т.е. независимо от другой пары признаков.
Сцепленное наследование
Г. Мендель проследил наследование семи пар признаков у гороха. Многие исследователи, повторяя опыты Менделя, подтвердили открытые им законы. Было признано, что эти законы носят всеобщий характер. Однако в 1906 г. английские генетики В.Бэтсон и Р.Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве. Потомки всегда повторяли признаки родительских форм. Стало ясно, что не для всех генов характерно независимое распределение в потомстве и свободное комбинирование.
Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков.
|
|
 Томас Гент Морган
(1886 — 1945)
Томас Гент Морган
(1886 — 1945)
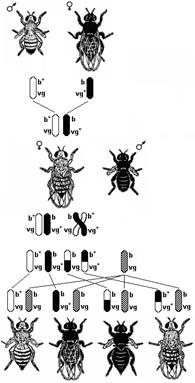 Рис. 327. Явление сцепленного
наследования признаков
у мушки дрозофилы.
Рис. 327. Явление сцепленного
наследования признаков
у мушки дрозофилы.
Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался выдающийся американский генетик Т. Морган. Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила. Мушка каждые две недели при температуре 25°С дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее.
Кроме того, они имеют всего 8 хромосом в диплоидном наборе и отличия по многочисленным признакам, могут размножаться в пробирках на дешевой питательной среде.
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибридов, имеющих серое тело и нормальные крыльяи (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев — над геном недоразвитых) (рис. 327). При проведении анализирующего скрещивания самки F1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% серые длиннокрылые и 41,5% черные с зачаточными крыльями) и лишь незначительная часть мушек имела перекомбинированные признаки (8,5% черные длиннокрылые и 8,5% серые с зачаточными крыльями).
Анализируя полученные результаты, Морган пришел к выводу, что гены, обусловливающие развитие серой окраски тела и длинных крыльев, локализованы в одной хромосоме, а гены, обусловливающие развитие черной окраски тела и зачаточных крыльев, — в другой. Явление совместного наследования признаков Морган назвал сцеплением. Материальной основой сцепления генов является хромосома. Гены, локализованные в одной хромосоме, наследуются совместно и образуют одну группу сцепления. Поскольку гомологичные хромосомы имеют одинаковый набор генов, количество групп сцепления равно гаплоидному набору хромосом (например, у человека 46 хромосом, или 23 пары гомологичных хромосом, соответственно количество групп сцепления в соматических клетках человека — 23). Явление совместного наследования генов, локализованных в одной хромосоме, называют сцепленным наследованием. Сцепленное наследование генов, локализованных в одной хромосоме, называют законом Моргана.
Вернемся к нашему примеру скрещивания мушек дрозофил. Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и ав, а отцовский — один тип — ав. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип ААВВ и аавв. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Аавв и ааВв. Каковы причины появления таких особей? Для объяснения этого факта необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Ав и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но поскольку кроссинговер происходит не во всех гаметах, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
В зависимости от особенностей образования гамет, различают:
©
|

©
|
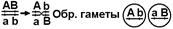
Соответственно этому различают:
© рекомбинантные (кроссоверные) особи — особи, возникшие с участием кроссоверных гамет;
© нерекомбинантные (некроссоверные) особи — особи, возникшие без участия кроссоверных гамет.
Гены в хромосомах имеют разную силу сцепления. Сцепление генов может быть:
© полным, если между генами, относящимися к одной группе сцепления, рекомбинация невозможна (у самцов дрозофилы полное сцепление генов, хотя у подавляющего большинства других видов кроссинговер протекает сходно как у самцов, так и у самок);
© неполным, если между генами, относящимися к одной группе сцепления, возможна рекомбинация.
Вероятность возникновения перекреста между генами зависит от их расположения в хромосоме: чем дальше друг от друга расположены гены, тем выше вероятность перекреста между ними. За единицу расстояния между генами, находящимися в одной хромосоме, принят 1 % кроссинговера. Его величина зависит от силы сцепления между генами и соответствует проценту рекомбинантных особей от общего числа потомков, полученных при скрещивании. Например, в рассмотренном выше анализирующем скрещивании получено 17% особей с перекомбинированными признаками. Следовательно, расстояние между генами серой окраски тела и длинных крыльев (а также черной окраски тела и зачаточных крыльев) равно 17%. В честь Т. Моргана единица расстояния между генами названа морганидой.
Результатом исследований Т.Моргана стало создание им хромосомной теории наследственности:
© гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов, причем набор генов каждой из негомологичных хромосом уникален;
© каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
© гены расположены в хромосомах в определенной линейной последовательности;
© гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
© сцепление генов может нарушаться в процессе кроссинговера; это приводит к образованию рекомбинатных хромосом; частота кроссинговера:
¨ является функцией расстояния между генами: чем больше расстояние, тем больше величина кроссинговера (прямая зависимость);
¨ зависит от силы сцепления между генами: чем сильнее сцеплены гены, тем меньше величина кроссинговера (обратная зависимость);
© каждый вид имеет характерный только для него набор хромосом — кариотип.
40.4. Генетика пола
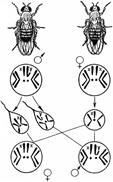 Рис. 328. Хромосомное определение пола у дрозофилы.
Рис. 328. Хромосомное определение пола у дрозофилы.
Как известно, большинство животных и двудомных растений являются раздельнополыми организмами, причем внутри вида количество особей мужского пола приблизительно равно количеству особей женского пола.
Пол можно рассматривать как один из признаков организма. Наследование признаков организма, как правило, определяется генами. Механизм же определения пола имеет иной характер — хромосомный (рис. 328).
Пол чаще всего определяется в момент оплодотворения. У человека женский пол является гомогаметным, то есть все яйцеклетки несут Х-хромосому. Мужской организм — гетерогаметен, то есть образует два типа гамет — 50% гамет несет Х-хромосому и 50% — Y-хромосому. Если
образуется зигота, несущая две Х-хромосомы, то из нее будет формироваться женский организм, если Х-хромосому и Y-хромосому — мужской.
|
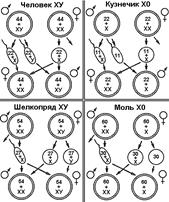 Рис. 329. Основные типы хромосомного определения пола.
Рис. 329. Основные типы хромосомного определения пола.
Из приведенной схемы видно, как происходит формирование в равных количествах двух групп особей, отличающихся набором половых хромосом.
Существует четыре основных типа хромосомного определения пола (рис. 329):
© мужской пол гетерогаметен; 50% гамет несут Х-, 50% —У-хромосому;
© мужской пол гетерогаметен; 50% гамет несут Х-, 50% —не имеют половой хромосомы;
© женский пол гетерогаметен; 50% гамет несут Х-, 50% —У-хромосому;
© женский пол гетерогаметен; 50% гамет несут Х-, 50% — не имеют половой хромосомы.
40.5. Наследование признаков,
сцепленных с полом
Генетические исследования установили, что половые хромосомы отвечают не только за определение пола организма — они, как и аутосомы, содержат гены, контролирующие развитие определенных признаков.
Наследование признаков, гены которых локализованы в Х- или Y-хромосомах, называют наследованием, сцепленным с полом.
Изучением наследования генов, локализованных в половых хромосомах, занимался Т.Морган.
У дрозофилы красный цвет глаз доминирует над белым. Проводя реципрокное скрещивание[41], Т.Морган получил весьма интересные результаты. При скрещивании красноглазых самок с белоглазыми самцами, в первом поколении все потомство оказывалось красноглазым. Если скрестить между собой гибридов F1, то во втором поколении все самки оказываются красноглазыми, а у самцов происходит расщепление — 50% белоглазых и 50% красноглазых. Если же скрестить между собой белоглазых самок и красноглазых самцов, то в первом поколении все самки оказываются красноглазыми, а самцы белоглазыми. В F2 половина самок и самцов — красноглазые, половина — белоглазые.
Объяснить полученные результаты наблюдаемого расщепления по окраске глаз Т.Морган смог, только предположив, что ген, отвечающий за окраску глаз, локализован в Х-хромосоме, а Y-хромосома таких генов не содержит.
Таким образом, благодаря проведенным скрещиваниям, был сделан очень важный вывод: ген цвета глаз сцеплен с полом, то есть находится в Х-хромосоме.
|
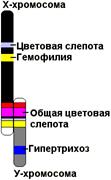 Рис. 330. Схема половых хромосом человека и сцепленных с ними генов.
Рис. 330. Схема половых хромосом человека и сцепленных с ними генов.
Х-хромосома человека содержит ряд генов, рецессивные аллели которых определяют развитие тяжелых аномалий (гемофилия, дальтонизм). Эти аномалии чаще встречаются у мужчин (так как они гетерогаметны), хотя носителем этих аномалий чаще бывает женщина.
У большинства организмов генетически активна только Х-хромосома, в то время как Y-хромосома практически инертна, так как не содержит генов, определяющих признаков организма. У человека лишь некоторые гены, не являющиеся жизненно важными, локализованы в Y-хромосоме (например, гипертрихоз — повышенная волосатость ушной раковины). Гены, локализованные в Y-хромосоме, наследуются особым образом — только от отца к сыну.
Полное сцепление с полом наблюдается лишь в том случае, если Y-хромосома генетически инертна. Если же в Y-хромосоме имеются гены, аллельные генам Х-хромосомы, характер наследования признаков иной. Например, если мать имеет рецессивные гены, а отец доминантные, то все потомки первого поколения будут гетерозиготны с доминантным проявлением признака. В следующем поколении получится обычное расщепление 3:1, причем с рецессивными признаками будут только девочки. Такой тип наследования называют частично сцепленным с полом. Так наследуются некоторые признаки человека (общая цветовая слепота, кожный рак).
40.6. Генотип целостная,
исторически сложившаяся система генов.
Изучая закономерности наследования, Г.Мендель исходил из предположения, что один ген отвечает за развитие только одного признака. Например, ген, отвечающий за развитие окраски семян гороха, не влияет на форму семян. Причем эти гены располагаются в разных хромосомах, и их наследование независимо друг от друга. Поэтому может сложиться впечатление, что генотип представляет собой простую совокупность генов организма. Однако сам Мендель в ряде опытов столкнулся с явлениями наследования, которые не могли быть объяснены с помощью открытых им закономерностей. Так, при изучении наследования окраски семенной кожуры, Мендель обнаружил, что ген, вызывающий образование бурой семенной кожуры, способствует также развитию пигмента и в других частях растения. Растения с бурой семенной кожурой имели цветки фиолетовой окраски, а растения с белой семенной кожурой — белые цветки. В других опытах, проводя скрещивание белой и пурпурной фасоли, он получил во втором поколении целый ряд оттенков — от пурпурного до белого. Мендель пришел к заключению, что наследование пурпурного цвета зависит не от одного, а от нескольких генов, каждый из которых дает промежуточную окраску. Можно говорить о том, что Мендель не только установил законы независимого наследования пар аллелей, но и заложил основы учения о взаимодействии генов.
После переоткрытия законов наследования признаков, многочисленные опыты подтвердили правильность установленных Менделем закономерностей. Вместе с тем, постепенно накапливались и факты, показывающие, что полученные Менделем числовые соотношения при расщеплении гибридного поколения не всегда соблюдались. Это указывало на то, что взаимоотношения между генами и признаками носят более сложный характер. Выяснилось, что:
© один и тот же ген может оказывать влияние на развитие нескольких признаков;
© один и тот же признак может развиваться под влиянием многих генов.
Как правило, взаимодействие генов имеет биохимическую природу, то есть оно основано на взаимодействии белков, синтезируемых под действием определенных генов. Взаимодействовать друг с другом могут как аллельные, так и неаллельные гены.
Дата: 2018-11-18, просмотров: 545.
Материалы представленные на сайте исключительно с целью ознакомления пользователям Интернета и не преследуют коммерческих целей или нарушение авторских прав.
© 2018 - 2026
|