Первое, второе и третье начала термодинамики. Определение понятия «температура».
Классическая термодинамика (“движение тепла”) – наука о взаимопревращениях различных видов энергии (главным образом теплоты и работы). Термодинамическая система – совокупность тел, способных энергетически взаимодействовать между собой и с другими телами и обмениваться с ними веществом и энергией.
Термодинамические параметры – величины, характеризующие состояние термодинамической системы (давление P, температура T, объем V и др.)
Различные термодинамические системы: изолированная (не обменивается энергией и веществом с другими системами), открытая (обменивается энергией и веществом), закрытая (обменивается только энергией), адиабатная (не обменивается теплом).
При постоянных внешних условиях любая термодинамическая система со временем приходит в равновесное состояние, характеризующееся неизменностью во времени термодинамических параметров и отсутствием в системе потоков вещества и энергии.
Термодинамическийпроцесс – изменение состояния системы, характеризующееся изменением ее термодинамических параметров.
Равновесный процесс – процесс, рассматриваемый как непрерывный ряд равновесных состояний системы.
Обратимый термодинамический процесс – процесс, после которого система и окружающая среда могут возвратиться в начальное состояние. Равновесный процесс всегда обратим, а обратимый процесс всегда протекает равновесным путем.
Различные термодинамические процессы: адиабатический (без обмена теплотой с внешней средой), изохорный (V = const), изобарный (P = const), изотермический (T = const).
Функция состояния – функция, которая определяется только параметрами системы и не зависит от пути перехода системы в то или иное состояние. Например, функция состояния идеального газа:
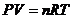
где P – давление, Па; V – объем, м3; n – число молей; R =8.31451 Дж/(моль K) – газовая постоянная; T – температура, K.
Первый закон термодинамики
Первый закон термодинамики - есть закон сохранения энергии: при любых физических взаимодействиях энергия не возникает и не исчезает, а только передается от одних тел другим или превращается из одной формы в другую.
Общая форма закона сохранения и превращения энергии имеет вид
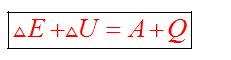

Но изучая тепловые процессы, мы будем рассматривать формулу 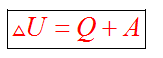
Согласно первому закону термодинамики, изменение внутренней энергии термодинамической системы при переходе из одного состояние в другое равно сумме работы, выполненной внешними силами, и количества теплоты, переданной системе извне
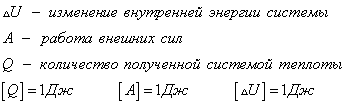
Сформулировать первый закон термодинамики можно иначе: количество теплоты, получаемое системой извне при ее переходе из одного состояния в другое, расходуется на повышение внутренней энергии системы и на работу, которую она выполняет против внешних сил
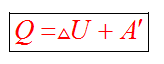
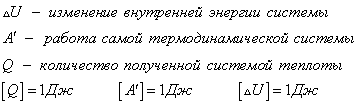
Например, вы кипятите чайник с водой. Количество тепла расходуется на их нагревание (увеличивается энергия частиц, то есть внутренняя энергия системы), а затем происходит приподнимание крышки - это работа, которую выполняет система.
Внешняя работа над системой равна работе системы, но с противоположным знаком
Обратный цикл
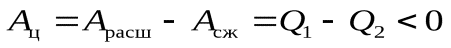
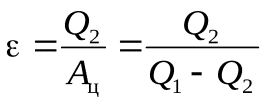 -холодильный коэф-нт
-холодильный коэф-нт
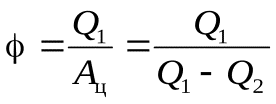 -отопительный коэф-нт
-отопительный коэф-нт
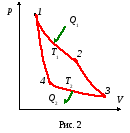 Цикл Карно– это цикл идеального двигателя, в котором тепло подводится и отводится в изотермических условиях при температурах нагревателяТ1и холодильникаТ2, переход отТ1кТ2и обратно осуществляется в адиабатных условиях.
Цикл Карно– это цикл идеального двигателя, в котором тепло подводится и отводится в изотермических условиях при температурах нагревателяТ1и холодильникаТ2, переход отТ1кТ2и обратно осуществляется в адиабатных условиях.
Ац = А12 + А23 + А34 + А41 (1)
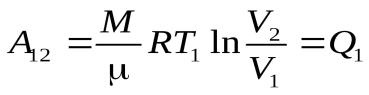 , (2)
, (2)
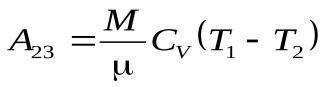 , (3)
, (3)
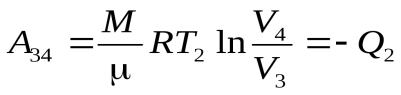 , (4)
, (4)
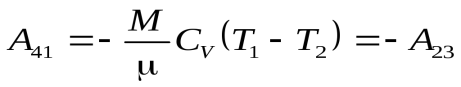 . (5)
. (5)
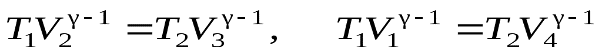
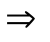
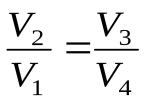 . (6)
. (6)
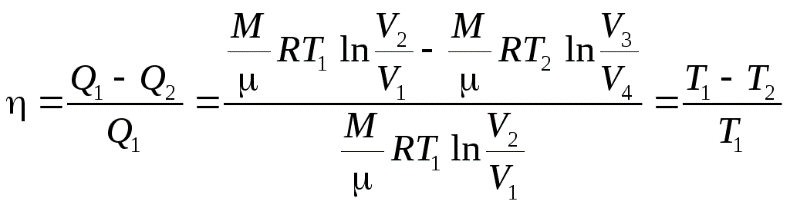
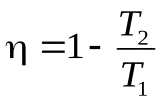 (7)
(7)
Теоремы Карно:
- Коэффициент полезного действия тепловой машины, работающей при данных значениях температур нагревателя и холодильника, не может быть больше, чем коэффициент полезного действия машины, работающей по обратимому циклу Карно при тех же значениях температур нагревателя и холодильника.
- Коэффициент полезного действия тепловой машины, работающей по циклу Карно, не зависит от рода рабочего тела, а зависит только от температур нагревателя и холодильника.
теорема Карно послужила основанием для установления термодинамической шкалы температур, такая термодинамическая шкала не связана со свойствами какого-то определенного термометрического тела.
Адиабатный процесс.
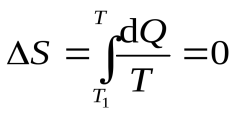
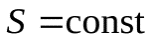 -процесс изоэнтропный(13)
-процесс изоэнтропный(13)
Изохорный процесс.
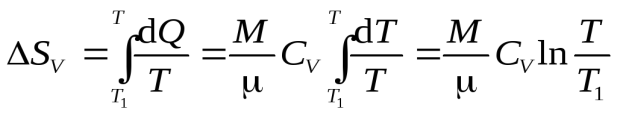






 Второй закон термодинамики устанавливаетнаправлениепротекания тепловых процессов.
Второй закон термодинамики устанавливаетнаправлениепротекания тепловых процессов.
Формулировка немецкого физика Р. Клаузиуса: невозможен процесс, единственным результатом которого была бы передача энергии путем теплообмена от тела с низкой температурой к телу с более высокой температурой.
Формулировка английского физика У. Кельвина: в циклически действующей тепловой машине невозможен процесс, единственным результатом которого было бы преобразование в механическую работу всего количества теплоты, полученного от единственного теплового резервуара.
Вероятностная формулировка австрийского физика Л.Больцмана: Он предложил рассматривать энтропию как меру статистического беспорядка замкнутой термодинамической системе. Всякое состояние системы c большим беспорядком характеризуется большим беспорядком. Термодинамическая вероятность W состояния системы – это число способов, которыми может быть реализовано данное состояние макроскопической системы, или число микросостояний, осуществляющих данное макросостояние. По определению термодинамическая вероятность W >> 1.
S = k ln W, (14)
где k = 1,38·10–23 Дж/К – постоянная Больцмана.
Таким образом, энтропия определяется логарифмом числа микросостояний, с помощью которых может быть реализовано макросостояние. Следовательно, энтропия может рассматриваться как мера вероятности состояния термодинамической системы.
Все самопроизвольно протекающие процессы в замкнутой системе, приближающие систему к состоянию равновесия и сопровождающиеся ростом энтропии, направлены в сторону увеличения вероятности состояния.
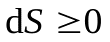 (15)
(15)
т.е. энтропия замкнутой системы может либо возрастать (в случае необратимых процессов), либо оставаться постоянной (в случае обратимых процессов).
Так как энтропия возрастает только в неравновесном процессе, то ее увеличение происходит до тех пор, пока система не достигнет равновесного состояния. Следовательно, равновесное состояние соответствует максимуму энтропии. С этой точки зрения энтропия является мерой близости системы к состоянию равновесия, т.е. к состоянию с минимальной потенциальной энергией.
Закон Гесса
Тепловой эффект химической реакции, проводимой в изобарно-изотермических или изохорно-изотермических условиях, зависит только от вида и состояния исходных веществ и продуктов реакции и не зависит от пути ее протекания:
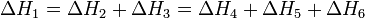
Следствия из закона Гесса:
1. Энтальпия реакции равна разности сумм энтальпий образования конечных и начальных участников реакций с учетом их стехиометрических коэффициентов.

2. Энтальпия реакции равна разности сумм энтальпий сгорания начальных и конечных реагентов с учетом их стехиометрических коэффициентов.
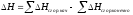
3. Энтальпия реакции равна разности сумм энергий связей Eсв исходных и конечных реагентов с учетом их стехиометрических коэффициентов.
В ходе химической реакции энергия затрачивается на разрушение связей в исходных веществах (ΣEисх) и выделяется при образовании продуктов реакции (–ΣEпрод). Отсюда
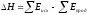
Следовательно, экзотермический эффект реакции свидетельствует о том, что образуются соединения с более прочными связями, чем исходные. В случае эндотермической реакции, наоборот, прочнее исходные вещества.
При определении энтальпии реакции по энергиям связей уравнение реакции пишут с помощью структурных формул для удобства определения числа и характера связей.
4. Энтальпия реакции образования вещества равна энтальпии реакции разложения его до исходных веществ с обратным знаком.
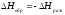
5. Энтальпия гидратации равна разности энтальпий растворения безводной соли 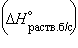 и кристаллогидрата
и кристаллогидрата 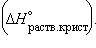
Из вышесказанного видно, что закон Гесса позволяет обращаться с термохимическими уравнениями как с алгебраическими, т. е. складывать и вычитать их, если термодинамические функции относятся к одинаковым условиям.
Закон Кирхгофа
Закон Кирхгофа гласит, что температурный коэффициент теплового эффекта химической реакции равен изменению теплоёмкости системы в ходе реакции. Уравнение Кирхгофа, являющееся следствием этого закона используется для расчёта тепловых эффектов при разных температурах.
Дифференциальная форма закона:
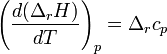
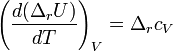
Интегральная форма закона:
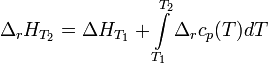
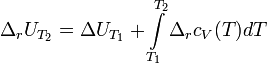
где 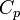 и
и 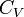 — изобарная и изохорная теплоёмкости,
— изобарная и изохорная теплоёмкости, 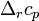 — разность изобарных
— разность изобарных
теплоёмкостей продуктов реакции и исходных веществ, 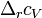 — разность изохорных
— разность изохорных
теплоёмкостей продуктов реакции и исходных веществ, а 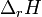 и
и 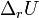 — соответствующие тепловые эффекты.
— соответствующие тепловые эффекты.
Теплоемкость-количество теплоты, необходимое для повышения температуры вещества на 1 градус.
Истинная теплоемкость- количество теплоты(бесконечно малое), которое необходимо для бесконечно малого изменения температуры.
Если разница 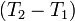 невелика, то можно принять
невелика, то можно принять 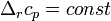 и
и 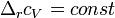 , соответственно интегральная форма уравнений примет следующий вид:
, соответственно интегральная форма уравнений примет следующий вид:
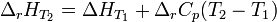
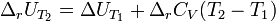
При большой разнице температур необходимо учитывать температурные зависимости теплоёмкостей: 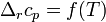 и
и 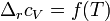
Для практических расчетов теплоемкость выражают в виде зависимости степенного ряда от температуры:
∆Cp=∆a+∆bT+∆cT2+∆c’T-2
Определение
Температура (от лат. temperatura — надлежащее смешение, нормальное состояние) — скалярная физическая величина, характеризующая состояние термодинамического равновесия макроскопической системы. Температура всех частей системы, находящейся в равновесии, одинакова. Если система не находится в равновесии, то между её частями, имеющими различную температуру, происходит теплопередача (переход энергии от более нагретых частей системы к менее нагретым), приводящая к выравниванию температур в системе.
Температура относится к интенсивным величинам, не зависящим от массы системы.
3.Термодинамика биологических систем. ''Жизнь с точки зрения физики" (Э.ШРЕДИНГЕР). Теорема Пригожина. Функция диссипации.
Любая живая система потребляет энергию и расходует её в виде работы и теплоты. С точки зрения термодинамики организму можно приписать свойства тепловой машины, подчиняющейся началам термодинамики. Первое начало термодинамики – закон сохранения энергии. Его применимость к живым системам надёжно доказана, но не выявляет никакого отличия их от неживых систем.
Второе начало термодинамики – закон энтропии– указывает, в каком направлении должны протекать естественные самопроизвольные процессы.В системемерой необратимости превращений энергии и одновременно мерой структурной неупорядоченности является энтропия. Согласно второму началу термодинамики энтропия изолированных систем не может уменьшаться, она либо возрастает, достигая максимума в состоянии термодинамического равновесия системы, либо, в крайнем случае, не изменяется. Эрвин Шредингер в книге “Что такое жизнь? С точки зрения физика” писал: “Если неживую систему изолировать или поместить в однородные условия, то всякое движение обычно очень скоро прекращается в результате различного рода трения, разность электрических или химических потенциалов выравнивается, вещества, которые имеют тенденцию образовывать химические соединения образуют их, температура выравнивается вследствие теплопроводности. Затем система в целом угасает, превращается в мёртвую инертную массу материи. Достигается состояние, при котором не происходит никаких заметных событий. Физик называет это стояние термодинамическим равновесием, или состоянием максимальной энтропии” [5]. Практически это состояние достигается быстро, но теоретически окончательное установление равновесия может происходить годами и столетиями. Очевидно, что всякий приток тепла извне увеличивает интенсивность теплового движения и повышает энтропию. Если мы будем расплавлять кристалл, то нарушим устойчивое расположение атомов или молекул, кристаллическая решётка превращается в непрерывно меняющееся случайное распределение атомов. Таким образом, согласно законам физики, материя стремится приблизиться к хаотическому состоянию.
Если рассматривать биологические процессы, то “… живая материя избегает перехода к равновесию”, структурная упорядоченность возрастает, а неупорядоченность, энтропия уменьшается. Например, при самосборке субклеточных структур или при эмбриональном развитии организма. Такие явления, как возникновение жизни, образование биосферы, прогрессивная эволюция, долгое время не вписывались в общую физическую картину мира, считались термодинамически маловероятными. Шредингер считал, что “деятельность живой материи, хотя и основана на законах физики, установленных к настоящему времени, но, по-видимому, подчиняется до сих пор неизвестным другим законам физики, которые, однако, как только они будут открыты, должны составить такую же неотъемлемую часть этой науки, как и первые”. В своих лекциях, прочитанных в 1943 г., он указывал, что организм избегает перехода к равновесию благодаря тому, что он питается и дышит, т.е. благодаря метаболизму. В процессе жизнедеятельности организм непрерывно увеличивает свою энтропию, приближаясь к опасному состоянию равновесия (максимальной энтропии), представляющему собой смерть. Избежать этого состояния организм может, только постоянно извлекая из окружающей среды “отрицательную энтропию” – то чем организм питается. Иными словами, живой организм постоянно привлекает на себя поток “отрицательной энтропии”, чтобы компенсировать этим увеличение энтропии, производимое в процессе жизни, и тем самым поддерживать себя постоянно на достаточно низком уровне энтропии. В метаболизме человек освобождается от всей той энтропии, которую он вынужден производить пока жив. “Отрицательная энтропия” или энтропия, взятая с отрицательным знаком – мера упорядоченности. Таким образом, с точки зрения Шредингера, живой организм непрерывно извлекает упорядоченность из окружающей среды. Например, высшие животные питаются крайне хорошо упорядоченным состоянием материи в более или менее сложных органических соединениях, после чего животные возвращают эти вещества в очень деградированной форме в окружающую среду. Для растений источником “отрицательной энтропии” является свет.
Книга Шредингера привлекла внимание физиков и химиков к проблемам биологии. Многие впервые высказанные им теоретические представления были подтверждены последующими исследованиями.
Реальные биологические системы в природе существенно открыты, гетерогенны, нелинейны, нестационарны и далеки от термодинамического равновесия. Совокупность этих свойств находится за пределами применимости второго начала термодинамики, даже с его новейшими расширениями. Сложно применять понятия энтропии и информации для описания общих свойств биологических систем.
Несмотря на это, современная термодинамика позволила дать новую трактовку фундаментальным биологическим процессам. Противоречие закону энтропии исчезает, если рассматривать организм в качестве единой системы вместе с окружающей средой, поскольку сам живой организм не является системой изолированной. Благодаря развитию термодинамики необратимых процессов к настоящему времени сформировалось представление, согласно которому по законам физики в открытых системах с потоком энергии вынужденно возникают динамические структуры в виде циклов, переносящих энергию, – упорядоченные круговороты вещества. При этом наиболее устойчивыми оказываются и поэтому «отбираются» функциональные структуры, состоящие из нескольких взаимодействующих циклов, т.е. более сложные динамические структуры, которые лучше вписываются в круговорот и эффективнее преобразуют проходящую через них энергию. Источником образования динамических структур на Земле служит поток солнечной энергии, который вызывает и организует круговороты в массах вещества – от простых физических (воды и воздуха) до сложных биологических.
Во многих случаях кажется, что они возникают сами по себе, и поэтому явление называют самоорганизацией структур. Появилось даже целое направление исследования таких процессов – синергетика(особенно важны в этом плане работы школы Ильи Пригожина, посвящённые проблеме возникновения организации из хаоса). Однако приложения синергетики к проблемам биологии дают лишь поверхностные аналогии, поскольку биологическая организация не исчерпывается структурной упорядоченностью.
Теорема Пригожина
Стационарным состоянием системы называют состояние, в котором процессы в системе так сбалансированы, что основные величины, характеризующие систему, остаются постоянными.
Живые организмы большую часть времени проводят именно в стационарном состоянии (точнее – переходя время от времени из одного стационарного состояния в другое, например – из состояния сна в состояние бодрствования и наоборот). Это свойство живых существ называют гомеостазом. Поддержание стационарного состояния (гомеостаз) имеет большое значение для нормальной жизнедеятельности. В организме всё время идут длинные цепи сопряжённых физических процессов и химических реакций. Эти процессы очень по разному зависят от температуры, рН и многих других параметров. Поэтому, если эти параметры меняются, нарушается согласование важных жизненных процессов. Чтобы такое не происходило, в организме существуют сложные системы регулирования, достаточно точно поддерживающие стационарное состояние. Наоборот, болезни в большинстве случаев сопровождаются нарушением гомеостаза.
Пригожин доказал положение, которое называется теоремой Пригожина: в стационарном состоянии продукция энтропии минимальна 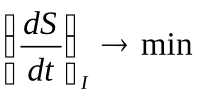 (в стационарном состоянии) (27).
(в стационарном состоянии) (27).
Нетрудно понять, что минимальная продукция энтропии соответствует минимальной диссипации свободной энергии (минимальным потерям энергии), то есть максимальному КПД. Поэтому можно сказать, что в стационарном состоянии КПД системы наибольший. Это тоже важное свойство стационарных состояний.
Выводы и заключение
Из материала лекции Вы усвоили, что законы термодинамики имеют большое значение для понимания самых разных сторон жизнедеятельности организма. Методы термодинамики успешно используются при исследовании самых разнообразных процессов, связанных с превращениями энергии в организме (питание, мышечное сокращение, обмен веществ, возникновение и проведение нервных импульсов, работа органов чувств и многое другое). Поэтому знание основных положений биологической термодинамики будет необходимо Вам при изучении большинства дисциплин.
Диссипативная функция
Изменение энтропии системы можно выразить через обобщенные силы и обобщенные координаты, характеризующие эту систему:
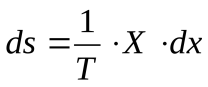
Отсюда можно найти скорость изменения энтропии (ds/dt). Эту величину называют диссипативной функцией:
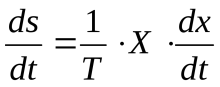
Например, при действии силы трения диссипативная функция выражается так:
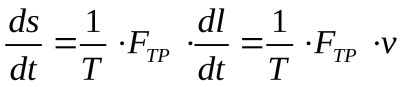
Если в системе идет химическая реакция, то обобщенной силой является химический потенциал (μx ), а обобщенной координатой— число молей вещества, вступающих в реакцию (v). Диссипативная функция имеет вид:
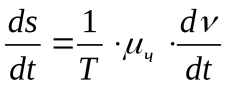
где dν/dt скорость реакции.
Если в системе происходит несколько процессов, то ее диссипативная функция определяется по формуле:
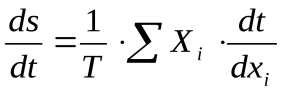
Диссипативную функцию, рассчитанную на единицу объема (V), называют удельной продукцией энтропии:
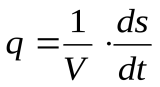
Ее можно считать количественной мерой необратимости процесса: чем больше q3, тем процесс дальше от обратимого.
Важным примером диссипации энергии является демпфирование. Этим термином обозначают уменьшение величины действующей силы (и, соответственно, ускорения) за счет перехода механической энергии в тепловую. В организме человека иживотных демпфирующими свойствами обладают скелетные мышцы, связочный аппарат, система кровообращения и др.Демпфирование играет важную роль в ослаблении нагрузок, действующих на организм. Это активный процесс, регулируемый центральной нервной системой. Выработка навыков демпфирования необходима летчикам, космонавтам, парашютистам,горнолыжникам — всем, кому приходится испытывать перегрузки. Следовательно, диссипация энергии не всегда должна рассматриваться как отрицательное явление. Примером использования диссипации энергии в лечебных целях служит высокочастотная электротерапия (например, УВЧ терапия), когда электромагнитная энергия в тканях организма диссипирует в тепло.
Диссипация энергии происходит во всех самопроизвольных процессах, идущих в изолированных системах. Так, при расширении газа его энтропия возрастает, поскольку с увеличением объема увеличивается число вариантов распределения молекул(растет термодинамическая вероятность). Возрастание энтропии свидетельствует об уменьшении свободной энергии.
Более сложным примером является смешение двух разных веществ. При образовании смеси работа не совершается, и может показаться, будто свободная энергия не изменяется. Однако в смеси веществ число вариантов распределения молекул больше, чем в сумме объемов, занимаемых ими до смешения. Следовательно, для смеси характерны более высокие величинытермодинамической вероятности и энтропии. Поэтому вещества довольно легко смешиваются, тогда как обратный процесс(самопроизвольное разделение смеси на компоненты) практически невероятен. Однако за счет подведения энергии извне можно добиться разделения смеси на составляющие ее вещества. Свободная энергия такой системы повышается.
В природе происходит немало процессов, вследствие которых свободная энергия той или иной системы возрастает, но они протекают только при наличии другой системы, «питающей» первую свободной энергией. Естественно, что при сопряжении процессов в двух системах свободная энергия одной из них обязательно понижается, причем общее изменение свободной энергии в полной системе, состоящей из нескольких взаимодействующих частей, всегда отрицательно, то есть ее энергия непременно претерпевает диссипацию.
Важную роль в процессах жизнедеятельности играет сопряжение электронных переходов в биологически важных молекулах с изменением их конформации (электронно-конформацион-ное взаимодействие — ЭКВ). Во многих процессах молекулы биополимеров (в первую очередь белков) так меняют свою третичную структуру, что их свободная энергия заметно возрастает. Этот запас свободной энергии может затем использоваться для совершения работы. Грубо говоря, молекула «работает» наподобие пружины: сжимается и разжимается. Подобное явление лежит в основе мышечного сокращения; так действуют многие ферменты. ЭКВ может происходить потому, что оно сопряжено с перестройкой электронной оболочки молекулы, то есть с переходом электронов на другие энергетические уровни, что приводит к освобождению заключенной в них энергии. Часть последней обеспечивает возрастание свободной энергии молекулы, испытавшей конформационную перестройку, а часть — диссипирует. Общее же изменение свободной энергии оказывается отрицательным в соответствии со вторым началом термодинамики, Сопряжение термодинамических процессов широко распространено в биологических системах.
Классификация артерий
По количественному соотношению эластических и мышечных элементов в стенке сосуда, артерии подразделяются на:
1. Артерии эластического типа.
2. Артерии смешанного типа (мышечно- эластического) типа.
3. Артерии мышечного типа.
Классификация вен
1. Вены безмышечного типа.
2. Вены мышечного типа, которые в свою очередь подразделяются на:
- вены со слабым развитием миоцитов
- вены со средним развитием миоцитов
- вены с сильным развитием миоцитов
Степень развития миоцитов зависти от локализации вены: в верхней части тела мышечный компонент развит слабо, в нижней – сильнее.
Работа и мощность сердца.
В соответствии с законом Пуазейля течение жидкости по сосудам определяется разностью давлений в начале и конце сосудов. В системе кровообращения этот перепад давлений обусловлен, в основном, работой сердца. Таким образом, по отношению к сосудистой системе сердце можно считать некоторым насосом. В отличии от обычного поршневого насоса, где площадь поршня постоянна, при работе сердца его рабочая поверхность - внутренняя поверхность желудочков - изменяется в различные фазы их сокращения. Сила F, действующая на кровь в желудочках, зависит от давления Р в их полости и площади внутренней поверхности S : F = P S. В начале систолы давление меньше, а объем и площадь поверхности полости желудочков - больше. В конце систолы давление возрастает, а внутренняя поверхность уменьшается. Количественное рассмотрение этих процессов и непосредственные физиологические эксперименты показывают, что при уменьшении объема внутренних полостей сердце развивает меньшую силу. Согласно закону Старлинга, сила, развиваемая при сокращении сердца, пропорциональна начальной длине волокон миокарда.
Не занимаясь рассмотрением сложных явлений, происходящих при сокращении сердца, рассчитаем его работу за одно сокращение, исходя из общих физических соображений. Эта работа А складывается из работы левого Ал и правого Апжелудочков:
А = Ал + Ап .
Работа левого желудочка, выбрасывающего кровь в большой круг кровообращения, естественно, больше чем работа правого желудочка. Считается, что Ап = 0,2 Ал . Таким образом А = 1,2 Ал.
Работа левого желудочка при выбросе систолического (ударного) объема крови в аорту затрачивается на преодоление сил давления крови в сосудистой системе и на сообщение крови кинетической энергии. Первая составляющая этой работы называется статическим компонентом, а вторая - кинетическим компонентом.
Статический компонент рассчитаем по формуле:
Аст= Рср Vс ,
где Рср - давление крови в аорте, Vс - систолический объем. Считая для человека в покое Рср = 100 мм.рт.ст. = 13,3 кПа и Vс= 60 мл = 6×10-5 м3, найдем: Аст » 0,8 Дж.
Кинетический компонент может быть рассчитан из формулы для кинетической энергии:
Ак = 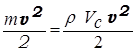 ,
,
где m - масса крови в ее систолическом объеме, r - плотность крови, v - линейная скорость крови при выбросе в аорту. Приняв r = 1,05×103кг/м3, v = 0,5 м/с, получим, что Ак = 0,008 Дж.
Таким образом, работа сердца за одно сокращение, определяемая по формуле:
А = 1,2 (PVc + 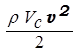 ) (32)
) (32)
составляет в покое примерно 1 Дж. Считая, что, в среднем, сердце сокращается один раз в секунду, определим работу сердца за сутки. Она составляет внушительную величину: 86400 Дж. Для сравнения, такую работу надо совершить, чтобы поднять груз массой в 1т на высоту около 9м.
Учитывая, что продолжительность систолы составляет около 0,3 с, и разделив работу сердца за одно сокращение на это время, получим для средней мощности сердца в покое значение 3,3 Вт.
При физической нагрузке возрастает систолический и минутный объем крови, увеличивается и скорость течения крови в аорте. Работа сердца резко увеличивается. При этом изменяется и соотношение между статическим и кинетическим компонентами. Как видно из приведенных численных значений, в покое доля кинетического компонента составляет около 1% от общей работы сердца. С повышением нагрузки растут оба слагаемых формулы (32), но второе слагаемое растет быстрее, так как величина кинетического компонента работы сердца пропорциональна квадрату увеличивающейся линейной скорости кровотока. Доля кинетического компонента в общей работе сердца возрастает и может достигать 30%.
Пульсовые колебания скорости кровотока. Пульсовые колебания давления (систолическое, диастолическое и среднее артериальное давление крови). Пульсовая волна. Уравнение для гармонической пульсовой волны. Формула скорости пульсовой волны.
Пульсовые колебания кровотока. При сокращении сердца кровь поступает из левого желудочка в восходящую аорту только во время периода изгнания. В ходе так называемых пульсовых колебаний скорость кровотока меняется следующим образом: сначала, после открытия аортальных клапанов, она резко возрастает, достигая максимума примерно
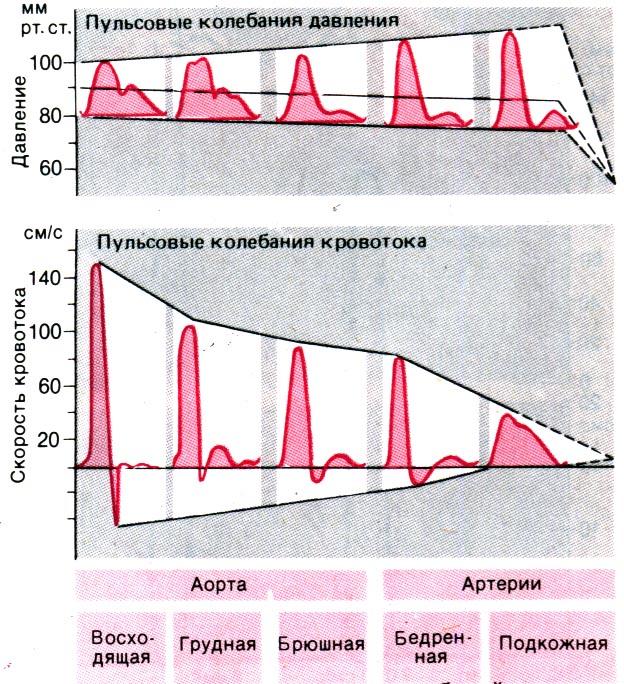
Рис. 20.9. Изменения пульсовых колебаний давления и кровотока в аорте и артериях ног. Следует отметить, что по мере удаления от сердца появляется антероградный кровоток во время диастолы и повышается систолическое давление (по Макдональду [15])
к концу первой трети периода изгнания; затем, к моменту окончания этого периода, она падает до нуля (рис. 20.9). От начала периода расслабления и до закрытия аортальных клапанов наблюдается кратковременный обратный ток крови в левый желудочек. Во время диастолы, до начала следующего периода изгнания, кровь в восходящей аорте обычно не движется.
У человека в состоянии покоя максимальная скорость кровотока в аорте (при длительности периода изгнания 0,25 с и величине ударного объема 70–90 мл) значительно превосходит 100 см/с; средняя же скорость в течение всего периода изгнаниясоставляет около 70 см/с. Во время большей части периода изгнания число Рейнольдса превосходит критическое значение, и кровоток в аорте носит турбулентный характер. По мере удаления от сердца амплитуда волны кровотока в аорте и крупных артериях постепенно снижается (в отличие от амплитуды волны
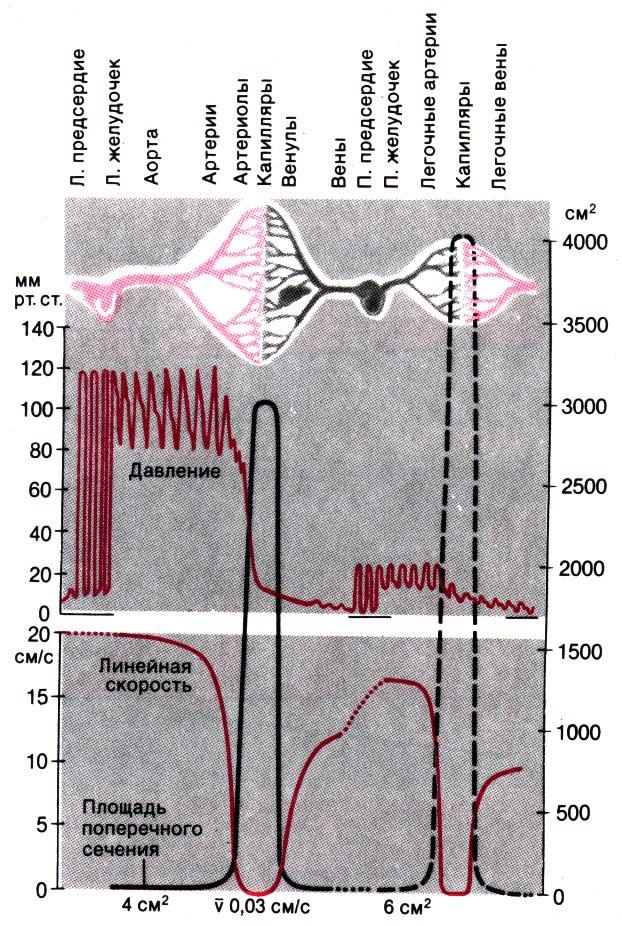
Рис. 20.10. Схема соотношения между площадью поперечного сечения, давлением и средней линейной скоростью кровотока в различных отделах сердечно–сосудистой системы
давления). В то же время в грудной аорте и периферических артериях во время диастолы наблюдается антероградный кровоток (рис. 20.9). Кратковременный обратный кровоток в начале периода расслабления в покое можно зарегистрировать даже в бедренной (или плечевой) артерии. Однако при увеличении сердечного выброса возрастает и скорость кровотока, и в конечном счете кривая записи пульса перестает пересекать нулевую линию.
В области концевых разветвлений артерий и артериол пульсирующий кровоток постепенно сменяется непрерывным. Однако при максимальном расширении сосудов небольшие колебания кровотока наблюдаются даже в капиллярах и мелких венах.
Скорость кровотока и площадь поперечного сечения сосудов. Поскольку кровоток, особенно в аорте и крупных артериях, носит пульсирующий характер, средняя скорость тока крови в этих сосудах значительно ниже, чем скорость во время систолы. Ее можно вычислить по формуле v = V/(πr2). В покое при сердечном выбросе, равном 96 мл/с, средняя линейная скорость кровотока в аорте радиусом 12–13 мм колеблется соответственно от 21,2 до 18,1 см/с (т.е. в среднем около 20 см/с; см. рис. 20.10 и табл. 20.4). При увеличении сердечного выброса средняя линейная скорость кровотока может превышать 100 см/с.
В связи с тем что средняя скорость кровотока обратно пропорциональна поперечному сечению сосудов, она значительно ниже в периферических артериях, особенно в концевых артериях и артериолах. Медленнее всего кровь течет в капиллярах: линейная скорость кровотока в них составляет 0,03 см/с (см. рис. 20.10 и табл. 20.4). Время прохождения крови через капилляр средней длины (около 750 мкм) составляет примерно 2,5 с.
Эквивалентная электрическая модель сердечно-сосудистой системы. Дипольный генератор электрического поля. Уравнение для потенциала электрического поля дипольного генератора на поверхности объёмного проводника. Схема электрического поля сердца
Требуется построить математическую модель такого процесса. Модель можно будет использовать для изучения работы сердца и системы кровообращения. Варьируя характеристики (эластичности артерий и вен, оббьем расширения и сжатия сердца, периферийного сопротивления, работы клапанов) можно проследить, как они влияют на работу сердца, например, на перепады давления, которые легко потом измерить.
Для построения модели целесообразно гидродинамическую систему заменить на эквивалентную электрическую схему. Установим аналогию между характеристиками исследуемой системы и параметрами электрической цепи.
Кровяное давление – потенциал (напряжение) U.
Объем крови - заряд q.
Поток крови –ток I.
Сопротивление периферийному кровообращению – сопротивление R.
Эластичность артерий и вен (способность расширяться и накапливать кровь) – емкости 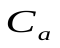 и
и 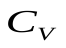 .
.
С 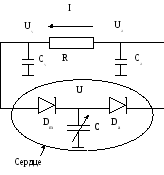 ердце моделируется с помощью диодовDa , Dm , соответствующих аортальному и митральному клапанам, и переменной емкости
ердце моделируется с помощью диодовDa , Dm , соответствующих аортальному и митральному клапанам, и переменной емкости 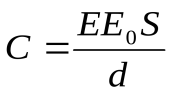 . Если раздвинуть пластинки конденсатора, емкость уменьшится. Заряд остается постоянным, поэтому напряжение
. Если раздвинуть пластинки конденсатора, емкость уменьшится. Заряд остается постоянным, поэтому напряжение 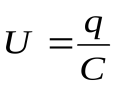 возрастает:d
возрастает:d  C
C  U .
U .
Пусть U, Ua , Uv - напряжения на конденсаторах C, Ca , Cv, т.е. U, Ua, Uv соответствуют давлениям в сердце, артериях и венах.
Эквивалентная электрическая схема:
Суммарный объем крови (Q) в сердце ( 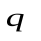 ), артериях (
), артериях ( 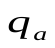 ) и венах (
) и венах ( 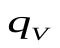 ) остается постоянным
) остается постоянным 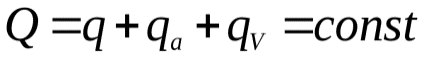 .Это можно использовать для задания начальных условий (в данном случае остается постоянным не напряжение, а заряд, так как емкость меняется скачком).
.Это можно использовать для задания начальных условий (в данном случае остается постоянным не напряжение, а заряд, так как емкость меняется скачком).
В момент t=0 происходит расширение стенки сосуда сердца. Будем имитировать эквивалентный процесс увеличением емкости конденсаторов. Пластинки конденсаторов сдвигаются, т.е. уменьшается d, емкость возрастает, а напряжение (и соответственно давление в сердце) падает d  C
C  U .
U .
Напряжение U становится меньше напряжения 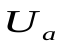 . ДиодDa закрывается, диод Dm открывается. Напряжения на конденсаторах
. ДиодDa закрывается, диод Dm открывается. Напряжения на конденсаторах 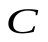 и
и 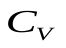 выравниваются,
выравниваются, 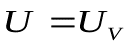 .(в электрической схеме они оказываются соединенными «на коротко»).
.(в электрической схеме они оказываются соединенными «на коротко»).
1. П 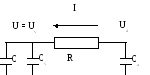 ри0<t<T/2 ¾ диастола. Электрическая схема при диастоле
ри0<t<T/2 ¾ диастола. Электрическая схема при диастоле
Так как 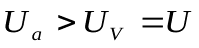 , идет разряд конденсатораCa и заряд конденсаторов С и Сv., в цепи протекает ток I.
, идет разряд конденсатораCa и заряд конденсаторов С и Сv., в цепи протекает ток I.
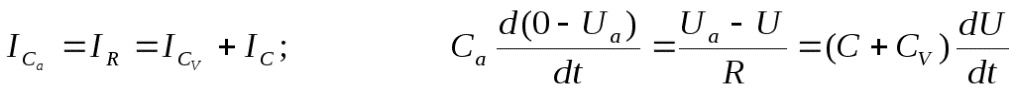
При t=T/2 емкость С резко уменьшается (например, раздвигаются пластины конденсатора, d увеличивается, падает). Напряжение U начинает резко расти: (d C U ). Этот момент соответствует максимальному пику (резкий всплеск давления в сердце и в артериях во время сжатия сердца).
Диод Dm закрывается. Как только U достигает значения 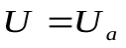 , диодDa открывается. Происходит перераспределение заряда между конденсаторами С и , но заряд на
, диодDa открывается. Происходит перераспределение заряда между конденсаторами С и , но заряд на 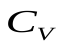 и напряжение
и напряжение 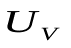 остаются постоянными во время скачка.
остаются постоянными во время скачка.
2. П 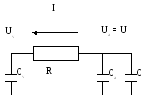 риT/2<t<T – систола. Электрическая схема при систоле:
риT/2<t<T – систола. Электрическая схема при систоле:
Так как 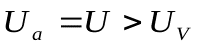 , идет разряд конденсаторов и С и заряд конденсатора .
, идет разряд конденсаторов и С и заряд конденсатора .

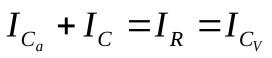 ;
; 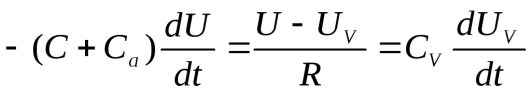 .
.
Далее периодический процесс повторяется.
Данные уравнения легко решаются и рассчитываются всеU и q: давление и объем крови в сердце, артериях и венах. При этом задается период, заряд Q, R,, Ca, CV, C (при систоле) и C (при диастоле). Графики изменения напряжения (давления) и заряда (объёма крови) на конденсаторах: С (в сердце), Са (в артериях) и Сv (в венах) показаны на рисунках.
Перепад артериального давления (среднее за систолу / среднее за диастолу) оказалось 112.5/64.3. Хотя напряжение и в условных единицах, это отношение приблизительно соответствует перепаду давления сердца взрослого человека.
В заключение следует отметить одно интересное обстоятельство. В электрической схеме постоянно протекает электрический ток. За счёт присутствия в схеме сопротивления постоянно происходит расход электрической энергии (W = I2Rt). Источника электрической энергии в схеме нет. Однако процесс носит периодический незатухающий характер. Может сложиться впечатление, что работа схемы напоминает вечный двигатель: происходит постоянный расход электрической энергии, в то время как извне энергия не поступает. Откуда берётся энергия?
На самом деле энергия поступает за счёт работы при изменении ёмкости конденсатора. Для того, чтобы перемещать пластины заряженного конденсатора, необходимо совершать работу против действия сил электрического поля. Таким образом, конденсатор с изменяющейся ёмкостью представляет собой источник, преобразующий механическую работу в электрическую энергию.
В реальной системе (сердце и система кровообращения) энергия поступает за счёт сжатия и расширения стенок сердца.
Таким образом, с помощью эквивалентной электрической схемы мы легко построили количественную модель и записали уравнение. Меняя эластичность вен и артерий, сопротивление периферийной системы и другие параметры можно проследить их влияние на изменения кровяного давления (которое легко измеряется).



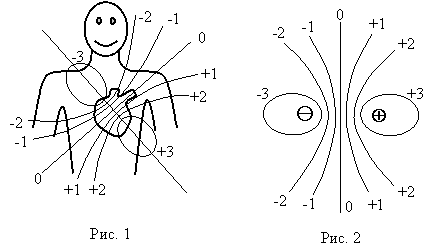
Источником электрического поля сердца являются электрические заряды – ионы, распределенные сложным образом в клетках и межклеточном пространстве миокарда. Картина эквипотенциальных линий электрического поля изображена на рис. 1 (в момент сокращения желудочков). Вид этих линий напоминает поле, создаваемое электрическим диполем (рис. 2).
Основной физической величиной для диполя является вектор электрического момента диполя 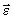 , равный по величине произведению
, равный по величине произведению
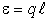 ,
,
|
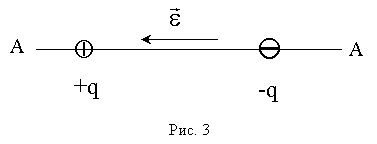
где 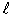 - расстояние между зарядами. При этом вектор направлен вдоль оси диполя АА от отрицательного заряда (-q) к положительному (+q) (рис. 3).
- расстояние между зарядами. При этом вектор направлен вдоль оси диполя АА от отрицательного заряда (-q) к положительному (+q) (рис. 3).
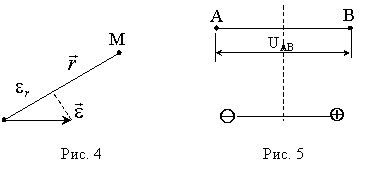
Электрическое поле диполя (рис. 2) в любой удаленной точке полностью определяется вектором . Так, значение потенциала j в некоторой точкеМ, удаленной на большое расстояние r от диполя,
|
зависит от проекции 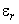 вектора на направление радиус-вектора точки М
вектора на направление радиус-вектора точки М 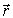 (рис. 4); k - коэффициент пропорциональности, зависящий от диэлектрической проницаемости среды, окружающей диполь, и от выбора системы единиц.
(рис. 4); k - коэффициент пропорциональности, зависящий от диэлектрической проницаемости среды, окружающей диполь, и от выбора системы единиц.
Из формулы 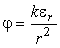 можно получить следующее важное для дальнейшего свойство электрического поля диполя: напряжение (разность потенциалов) UАВ между двумя равноудаленными от диполя точками (А и В, рис. 5) прямо пропорционально проекции e АВ вектора на направление АВ:
можно получить следующее важное для дальнейшего свойство электрического поля диполя: напряжение (разность потенциалов) UАВ между двумя равноудаленными от диполя точками (А и В, рис. 5) прямо пропорционально проекции e АВ вектора на направление АВ:
UАВ~ eАВ. (3)
Поляризацией вещества в электрическом поле называют явление, обусловленное упорядочением в этом веществе связанных электрических зарядов, которые ориентируются так, что образуемое ими дополнительное электрическое поле направлено в сторону, противоположную внешнему полю.
Поляризация в диэлектриках связана с существованием в диэлектрике или образованием в нем под действием электрического поля электрических диполей, т. е. систем, состоящих из двух равных, но противоположных по знаку зарядов q, находящихся на расстоянии l друг от друга.
Основная характеристика диполя – его дипольный момент.
Дипольный момент р – вектор, численно равный произведению величины заряда на длину диполя (p = ql) и направленный от отрицательного заряда к положительному.
Поляризация может быть вызвана несколькими качественно различными причинами.
Понятие мембранного транспорта и его биологическое значение. Виды мембранного транспорта и их особенности. Химический и электрохимический потенциал веществ. Уравнение для расчета химического и электрохимического потенциалов.
Важным элементом функционирования мембран является их способность пропускать или не пропускать молекулы (атомы) и ионы. Вероятность такого проникновения частиц зависит как от направления так и от разновидности молекул и ионов. Транспортным переносом частиц называют необратимые процессы в результате которых в физической системе происходит пространственное перемещение (перенос) массы, импульса, энергии, заряда или какой – либо другой физической величины.
Есть пассивный и активный транспорт веществ через биологические мембраны. Пассивный транспорт веществ осуществляется за счет энергии, сконцентрированной в каком-либо градиенте и энергии метаболических процессов клеток непосредственно на этот перенос не расходуется. Пассивный транспорт всегда происходит по направлению градиентов химического и электрохимического потенциалов, то есть от более высокого энергетического уровня к более низкому, результатом чего является уменьшение градиентов, если нет других процессов, которые обеспечивают их поддержание на постоянном уровне. Различают следующие типы пассивного транспорта веществ и ионов через биологические мембраны: 1. Простая диффузия. 2. Перенос через поры (каналы) по градиенту концентрации вещества или иона. 3. Облегченная диффузия. Активным транспортом называют процесс переноса веществ или ионов против их градиентов, который может осуществляться за счет энергии метаболического процесса. Следовательно этот процесс может протекать самопроизвольно и является сопряженным с поставляющим для него энергию процессом. Различают 2 разновидности активного транспорта: Под первично – активным транспортом понимают транспорт веществ против градиента их концентраций при непосредственном сопряжении с реакцией типа гидролиза АТФ. При вторично – активном транспорте используется энергия электрохимического градиента, либо какого – то вещества, для “накачивание” другого вещества против градиента его градиента.
Химическим потенциалом данного вещества mк называется величина, численно равная энергии Гиббса, приходящаяся на 1 моль этого вещества. Математически химический потенциал определяется как частная производная от энергии Гиббса G по количеству какого-либо вещества, при постоянстве температуры, давления и количестве всех других веществ m1 (l не = к): mк = (¶G/¶mк) Р, T, m (l не = к), для разбавленного раствора концентрации вещества С: m =m0 + RTlnC, где m0 – стандартный химический потенциал, численно равный химическому потенциалу данного вещества при его концентрации 1 моль/л в растворе. Электрохимический потенциал m - величина, численно равная, энергии Гиббса G на один моль данного вещества, помещенного в электрическом поле. Для разбавленных растворов: m = =m0 + RTlnC + ZFj, где F – 96500 кл/моль – число Фарадея, Z – заряд иона электролита, j - потенциал электрического поля, T [K] – температура.
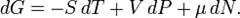
Химическим потенциалом данного вещества mк называется величина, численно равная энергии Гиббса, приходящаяся на один моль этого вещества. Математически химический потенциал определяется как частная производная от энергии Гиббса, G по количеству k-гo вещества, при постоянстве температуры Т, давления Р и количеств всех других веществ ml (l¹k).
mk = (¶G/¶mk )P,T,m
Для разбавленного раствора концентрации вещества С:
m = m0 + RTlnC
где m0- стандартный химический потенциал, численно равный химическому потенциалу данного вещества при его концентрации 1 моль/л в растворе.
Электрохимический потенциал m- величина, численно равная энергии Гиббса G на один моль данного вещества, помещенного в электрическом поле.
Для разбавленных растворов
m = mo + RTlnC + ZFj
где F = 96500 Кл/моль - число Фарадея, Z - заряд иона электролита (в элементарных единицах заряда), j - потенциал электрического поля, Т [К] – температура.
Пассивный транспорт ионов. Уравнение ТЕОРЕЛЛА, НЕРНСТА - ПЛАНКА.
Пассивный транспорт – это перенос вещества из мембраны с большим значением электрохимического потенциала к местам с его меньшим значением.
Пассивный транспорт:
1. Фильтрация.
2. Простая диффузия:
a). Осмос,
б). Через липидный слой,
в). Через поры в липидном слое,
г). Через белковую пору.
3. Облегченная диффузия:
а). С подвижным переносчиком,
б). С фиксированным переносчиком.
Пассивный транспорт идет с уменьшением энергии Гиббса и поэтому этот процесс может идти самопроизвольно без затраты энергии. Направление пассивного транспорта μ1 → μ2, Jm, μ1 больше μ2 . Плотность потока вещества Jm при пассивном транспорте подчиняется уравнению Теорелла:
Jm = - UCd μ/dx,
где U – подвижность частиц, C – концентрация. Знак минус показывает, что перенос происходит в сторону убывания μ.
Плотность потока вещества – это величина, численно равная количеству вещества, переменного за единицу времени через единицу площади поверхности, перпендикулярной направлению переноса: Jm = m/St [моль/м2с]. Подставим в первое выражение для электрохимического потенциала, m = m0 + RTlnC + ZFj, получим для разбавленных растворов при m0 = постоянной уравнение Нернста – Планка: Jm = - URT dc/dx - UCFZ dj/dx. Итак, могут быть 2 причины переноса вещества при пассивном транспорте: градиент концентрации dc/dx и градиент электрохимического потенциала dj/dx. Знаки минус перед градиентами показывают, что градиент концентрации вызывает перенос вещества от места с большей концентрацией к местам с его меньшей концентрацией, а градиент электрохимического потенциала вызывает перенос положительных зарядов от мест с большим к местам с меньшим положительным. Фильтрацией называется движение раствора через поры в мембране под действием градиента давления. Скорость переноса при фильтрации подчиняется закону Пуазейля:
dV/dt = P1 – P2/W,
где dV/dt - объемная скорость переноса раствора, W – гидравлическое сопротивление, W = 8ήl/πr4, l – длина поры, r – ее радиус, ή - коэффициент вязкости раствора. Явление фильтрации играет важную роль в процессе переноса воды через стенки кровеносных сосудов
26. Активный транспорт ионов Na+ и К+. Структура и работа Nа+и К+-зависимой АТФазы.
Активный транспорт ионов – транспорт ионов против их электрохимического градиента. Наиболее хорошо изученным мембранным компонентом, реализует активный транспорт, является натриевая помпа, молекулярной основой является Na+ - K+ - активируемая, Mg – зависимая АТФаза (Na+ - K+ - АТФаза). Впервые активируемую ионами Na+ и K+ - АТФаза обнаружил в 1957г. Скоу в гомогенате периферических нервов краба, так же в почках, солевых железах, мозге, электрическом органе, эритроцитах и других. Na+ - K+ - АТФаза относится к ферментам, затрачивают энергию АТФ на осуществление конформационных превращений, конечным результатом является перенос 3 ионов Na+ и 2 ионов K+ через плазматическую мембрану против их градиентов (осуществляя первично – активный транспорт одновалентных катионов). Фермент на мембране электрохимического градиента одновалентных катионов, энергия используется для процессов возбуждения нервной и мышечной ткани, транспорта аминокислот. Сахаров и других метаболитов через плазматическую мембрану. Na+ - K+ - АТФаза является глобулярным белком, имеющим олигомерную природу с протяженной структурой в толще липидного бислоя. Состоит из 2 субъединиц: α – субъединица (содержит приблизительно 1012 аминокислотных остатков), β – субъединица (приблизительно 300). α – субъединица выполняет каталитическую функцию, а β – субъединица играет важную роль в образовании α, β – комплексов в мембране. Удаление β – субъединицы приводит к снижению активности фермента. Полипептидная цепь β – субъединицы пронизывает мембрану 1 раз, а α – субъединица – 10 раз. Центр связывания сердечных гликозидов – ингибиторов фермента – расположен на внешней стороне мембраны, здесь же расположен и центр связывания ионов K+ . Активация фермента натрием осуществляется изнутри клетки, где расположен и центр, гидролизующий АТФ. Na+ - K+ - АТФаза является электрогенным насосом, работающим по принципу конформационно – лабильной поры. Конфирмационные изменения переносчика обеспечивают транспорт 3 ионов Na+из клетки и 2 ионов K+ в клетку. Конформационные переходы молекулы фермента (и перенос ионов) нуждаются в источнике энергии. Гидролиз АТФ, Na+ - K+ - АТФаза обеспечивает энергетически активный транспорт ионов. Основные стадии реакционного цикла фермента:
1) В присутствие ионов Na+ фермент гидролизует АТФ и переносит терминальный фосфат на боковой радикал остатка аспарагиновой кислоты полипептидной цепи, образуя фосфофермент (E1P).
2) Фосфорилирование – фермент изменяет конформацию то есть, что ионы Na+ теряют способность высвобождаться как с внутренней, так и с внешней стороны мембраны (окклюдированные ионы).
3) Ионы Mg2+ активируют конфирмационный E1P – E2P переход, сопровождается пространственными перемещениями в липидном бислое отдельных частей белковой глобулы – центр связывания ионов Na+ становится открытым во внешнеклеточное пространство и более гидрофобным, отсюда следует, что сродство ионов Na+ к активному центру фермента уменьшается и они переходят в водную фазу (стадия переноса ионов Na+ через мембрану).
4) Активный центр фермента связывается с ионами K+, так же как и Na+ подвергается окклюзии.
5) Связывание ионов K+ с E2P формой фермента сопровождается конформационными изменениями фермента, что следует окружение фосфатной группы, связанной с глобулой белка, становится более гидрофильным и доступным для атаки молекулами воды. Неорганический фосфат высвобождается во внутриклеточную среду.
6) Дефосфорилирование фермента – Конформационные изменения глобулы белка, ведет к уменьшению сродства фермента к ионам K+ и они диссоциируют от центра связывания в цитоплазму. Их место занимает Na+. Эта стадия цикла одновременно подготавливает фермент к началу нового цикла: конформер E2 превращается в конформер E1. АТФ ускоряет этот конформационный переход, повышая сродство фермента к Na+ и уменьшая его сродство к K+ . Если градиенты концентрации ионов K+и Na+ увеличить до такой степени, что насос не сможет с ними справиться, то ионы будут проходить через мембрану по градиенту концентрации и фермент начнет работать не как АТФаза, а как АТФ – синтетаза, то есть фермент из фосфата и АДФ будет синтезировать АТФ. В плазмолемме канальцевого эпителия почки функционируют 2 типа натриевых насосов:
1). Na+ - K+ - АТФаза, обеспечивает антипорт K+ и Na+ .
2). Осуществляет симпорт Na+ и Cl-.
27. Значимость ионных градиентов, создаваемых системами активного транспорта, для жизнедеятельности клеток. Физические принципы вторично-активного транспорта. Транспорт аминокислот, сахаров. Na+- Са++-обмен.
При первично – активном транспорте молекулярной системы, интегрированные в мембраны, трансформируют энергию химических связей органических молекул в энергию электрохимических градиентов ионов. Этот вид свободной энергии, в свою очередь, может быть использован другими переносчиками для транспорта веществ или ионов против их концентрационных градиентов. Это вторично – активный транспорт. Na – Ca – обмен. Na – Ca – обменный переносчик может функционировать в двух режимах: 1). Вход трех Na+ в обмен на выход Ca2+. 2). При снижении электрохимического градиента – выход Na в обмен на Ca.
Транспорт глюкозы. Белок – переносчик использует для работы энергию электрохимического градиента ионов Na. Na проникает в клетку по электрохимическому градиенту, что осуществляет переносчик, транспортируя вместе с ним глюкозу. Чем выше электрохимический градиент Na+ , тем выше скорость транспорта глюкозы. Аминокислоты переносятся как глюкоза.
Для более глубокого понимания процессов вторично-активного транспорта следует рассмотреть гипотетическую схему, показывающую, как градиент концентрации Na+мог бы приводить в действие переносчик глюкозы. Переносчик глюкозы может находиться в двух альтернативных конформационных состояниях – пинг и понг. В состоянии понг активный центр переносчика раскрыт во внеклеточное пространство и ионы натрия могут занимать свои участки связывания. По мере насыщения натриевых центров увеличивается сродство центров связывания переносчика к переносимому против градиента концентации веществу. Насыщение центров связывания глюкозы вызывает конформационную перестройку белковой глобулы переносчика, в результате чего активный центр переносчика оказывается раскрыт во внутриклеточное пространство (состояние пинг). Поскольку концентрация ионов натрия во внутриклеточном пространстве значительно ниже натрий покидает белок-переносчик, в результате чего сродство переносчика к глюкозе уменьшается, и она оказывается внутри клетки.Освобождение переносчика от натрия и молекул глюкозы приводит к возвращению белковой глобулы переносчика в первоначальное конформационное состояние.
Типы ионных каналов
Описано более 100 разновидностей ионных каналов, для классификации которых используют различные подходы. Один из них основан на учете различий в строении каналов и в механизмах функционирования. При этом ионные каналы можно разделить на несколько типов:
§ пассивные ионные каналы, или каналы покоя;
§ каналы щелевых контактов;
§ каналы, состояние которых (открытое или закрытое) контролируется влиянием на их воротный механизм механических факторов (механочувствительные каналы), разности потенциалов на мембране (потенциалзависимые каналы) или лигандов, связывающихся с каналообразующим белком на внешней или внутренней стороне мембраны (лигандзависимые каналы).
Натриевые каналы
В клетках организма широко представлены потенциалзависимые, потенциалнезависимые (лигандзависимые, механочувствительные, пассивные и др.) натриевые каналы.
Калиевые каналы
Существуют потенциалзависимые и потенциалнечувствительные калиевые каналы. Среди последних выделяют пассивные, лигандзависимые и другие типы калиевых каналов. Как правило, калиевые каналы имеются в мембранах тех же клеток и тканей, в которых содержатся натриевые каналы. Одной из причин такого параллелизма в расположении этих ионных каналов является то, что ионы Na+ и К+ являются важнейшими катионами, от характера распределения и перемещения которых зависит возникновение и изменение электрических потенциалов как одной из важнейших форм передачи информационных сигналов в организме.
Существует целое суперсемейство калиевых ионных каналов, которые подразделяются но особенностям структуры, месту локализации и свойствам каналов на отдельные семейства, типы и подтипы. Описано более трех десятков калиевых каналов, и дать их подробную характеристику не представляется возможным. Поэтому в качестве примеров будут приведены описания тех семейств и типов ионных каналов, которые имеют отношение прежде всего к сигнальным путям и механизмам контроля нервных и мышечных процессов.
Зубец Р
§ Отображает последовательную деполяризацию правого+левого предсердий.
§ Должен быть положительным в I и II отведениях, двухфазным в V1.
§ Ширина Р должна быть меньше 0,12 сек, высота - меньше 0,25 мВ.
Интервал PQ
§ Определяется от начала зубца Р до начала комплекса QRS.
§ Интервал PQ соотвтствует времени сокращения предсердий + времени проведения через AV-узел.
§ В норме длительность PQ составляет 120-200 мсек (3-5 маленьких клеточек).
Зубец Q
§ Это первый отрицательный зубец комплекса QRS, предшествующий зубцу R.
§ В норме зубец Q отображает деполяризацию межжелудочковой перегородки.
§ Небольшой (до 2 мм) зубец Q может присутствовать в "левых" отведениях I, aVL, V5, V6.
§ Зубец Q более 2 мм может присутствовать в III отведении, но он должен исчезнуть, если записать ЭКГ попросив пациента глубоко вдохнуть, например:
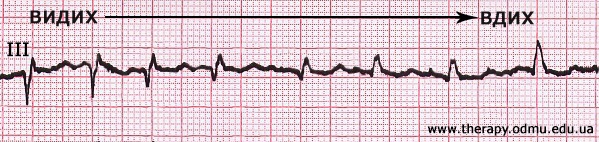
§ Патологическим считается зубец Q, превышающий 1/4 амплитуды зубца R, шириной 40 мсек и более, или если он обнаруживается в отведениях V1-V3.
Комплекс QRS
§ Отображает деполяризацию (сокращение) обоих желудочков (подробнее про QRS).
§ В норме ширина QRS составляет 0,07-0,1 сек. (2-2,5 маленьких клеточки).
§ Амплитуда QRS в стандартных отведениях должна быть выше 0,5 мВ (5 клеточек), в грудных - больше 1,0 мВ (10 клеточек).
Сегмент ST
§ Сегмент ST отображает то время, когда оба желудочка сокращены, и их реполяризация (расслабление) только начинается.
§ Сегмент ST измеряют в 60 мсек (полторы маленьких клетки) от точки J. В норме в отведениях V1-V3 может отмечаться элевация ST с максимумом в V2 до 0,25 mV, в других отведениях элевация в 0,1 mV считается патологической.
§ Читайте подробнее про все виды элевации и депрессии сегмента ST.
Зубец Т
§ Отображает процесс быстрой реполяризации (расслабления) желудочков.
§ Зубец Т конкордантен комплексу QRS (т.е. основная направленность Т совпадает с основной направленностью QRS)
§ Может быть отрицательным в отведениях aVL, III, V1; должен быть положительным в I, II, V3-V6.
§ В отведениях от конечностей - до 0,5 mV, в грудных отведениях - до 1,0 mV.
§ Подробно про зубец Т и его патологические изменения читайте в отдельной статье про зубец Т.
Зубец U
§ Отображает конечную стадию реполяризации желудочков, может отсутствовать на ЭКГ.
§ Сильнее всего выражен в отведениях V2-V3, где достигает 0,3 mV (обычно - около 10% высоты T)
§ Преимущественно проявляется при брадикардии.
§ Отрицательный или сливающийся с зубцом Т зубец U считается патологическим.
Понятие об электрокардиографических отведениях. Стандартные, усиленные и грудные электрокардиографические отведения. Требования, предъявляемые к электродам, используемым для регистрации биопотенциалов.
Стандартные отведения
Три стандартных отведения образуют равносторонний треугольник (треугольник Эйнтховена), вершинами которого являются правая и левая руки, а также левая нога с установленными на них электродами. Гипотетическая линия, соединяющая два электрода, участвующие в образовании электрокардиографического отведения, называется осью отведения. Осями стандартных отведений являются стороны треугольника Эйнтховена (рис. 1. 2).
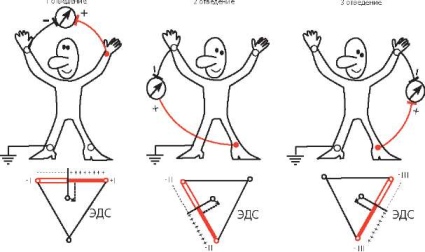
Рис. 1.2.
Особенности распространения возбуждения в сердечной мышце. Суммарный вектор ЭДС сердца. Электрическая ось сердца. Определение положения электрической оси сердца по результатам анализа ЭКГ, снятой в стандартных отведениях.
Возбудимость сердечной мышцы. Сердечная мышца менее возбудима, чем скелетная. Для возникновения возбуждения в сердечной мышце необходимо применить более сильный раздражитель, чем для скелетной. Установлено, что величина реакции сердечной мышцы не зависит от силы наносимых раздражений (электрических, механических, химических и т. д.). Сердечная мышца максимально сокращается и на пороговое, и на более сильное по величине раздражение.
Проводимость. Волны возбуждения проводятся по волокнам сердечной мышцы и так называемой специальной ткани сердца с неодинаковой скоростью. Возбуждение по волокнам мышц предсердий распространяется со скоростью 0,8—1,0 м/с, по волокнам мышц желудочков— 0,8—0,9 м/с, по специальной ткани сердца—2,0—4,2 м/с.
Сократимость. Сократимость сердечной мышцы имеет свои особенности. Первыми сокращаются мышцы предсердии, затем—папиллярные мышцы и субэндокардиальный слой мышц желудочков. В дальнейшем сокращение охватывает и внутренний слой желудочков, обеспечивая тем самым движение крови из полостей желудочков в аорту и легочный ствол.
Физиологическими особенностями сердечной мышцы является удлиненный рефрактерный период и автоматия. Теперь о них поподробнее.
Рефрактерный период. В сердце в отличие от других возбудимых тканей имеется значительно выраженный и удлиненный рефрактерный период. Он характеризуется резким снижением возбудимости ткани в течение ее активности. Выделяют абсолютный и относительный рефрактерный период (р.п.). Во время абсолютного р.п. какой бы силы не наносили раздражения на сердечную мышцу, она не отвечает на него возбуждением и сокращением. Он соответствует по времени систоле и началу диастолы предсердий и желудочков. Во время относительного р.п. возбудимость сердечной мышцы постепенно возвращается к исходному уровню. В этот период мышца может ответить на раздражитель сильнее порогового. Он обнаруживается во время диастолы предсердий и желудочков.
Сокращение миокарда продолжается около 0.3 с, по времени примерно совпадает с рефрактерной фазой. Следовательно, в период сокращения сердце неспособно реагировать на раздражители. Благодаря выраженному р.п. .р.рррр.п., который длится больше чем период систолы, сердечная мышца неспособна к тетаническому (длительному) сокращению и совершает свою работу по типу одиночного мышечного сокращения.
Автоматия сердца. Вне организма при определенных условиях сердце способно сокращаться и расслабляться, сохраняя правильный ритм. Следовательно, причина сокращений изолированного сердца лежит в нем самом. Способность сердца ритмически сокращаться под влиянием импульсов, возникающих в нем самом, носит название автоматии.
В сердце различают рабочую мускулатуру, представленную поперечнополосатой мышцей, и атипическую, или специальную, ткань, в которой возникает и проводится возбуждение.
У человека атипическая ткань состоит из:
синоаурикулярного узла, располагающегося на задней стенке правого предсердия у места впадения полых вен;
атриовентрикулярного (предсердно-желудочкого) узла находящегося в правом предсердии вблизи перегородки между предсердиями и желудочками;
пучка Гиса (председно-желудочковый пучок), отходящего от атриовентрикулярного узла одним стволом. Пучок Гиса, пройдя через перегородку между предсердиями и желудочками, делится на две ножки, идущие к правому и левому желудочкам. Заканчивается пучок Гиса в толще мышц волокнами Пуркинье. Пучок Гиса—это единственный мышечный мостик, соединяющий предсердия с желудочками.
Суммарный вектор ЭДС сердца
Для определения по зубцам ЭКГ направления ЭДС сердца нужно помнить, что зубец, расположенный над изоэлектрической линией, обусловлен направлением ЭДС сердца в продолжение соответствующего интервала времени в сторону положительного полюса данного отведения.
Зубец, направленный вниз от изоэлектрической линии, свидетельствует об ориентации ЭДС в сторону отрицательного полюса данного отведения.
Учитывая, что направление и полярность осей отведения известны, определить направление и величину суммарной ЭДС сердца в любой момент возбуждения нетрудно.
Суммарный вектор ЭДС сердца в норме во время деполяризации направлен влево и вниз, в связи с чем суммарный вектор возбуждения проецируется в основном на положительные части осей отведений во фронтальной плоскости.
Это приводит к тому, что доминирующим зубцом в этих отведениях будет положительный зубец (зубец R), Исключением является отведение aVR.
Суммарный вектор ЭДС сердца проецируется на отрицательную часть этого отведения, и доминирующим зубцом будет отрицательный зубец.
По зубцам стандартных и однополюсных отведений от конечностей можно найти направление суммарного вектора ЭДС сердца во фронтальной плоскости. В практической электрокардиографии это положение применяется для определения направления электрической оси сердца.
Аналогичным образом используются оси грудных отведений для изучения векторов ЭДС в горизонтальной плоскости.
Изменения зубцов ЭКГ в определенных отведениях при помощи известного расположения осей этих отведений позволяют определить отклонения в направлении и величине вектора ЭДС сердца, которые связаны обычно с преобладанием ЭДС какого-либо отдела сердца.
ЭОС - это суммарное направление электрической волны, которая проходит по желудочкам в момент сокращения. Следует понимать, что электрическая ось сердца не является его анатомической осью. Более того, очень часто при гипертрофии левого или правого желудочков ЭОС в соответствующую сторону отклоняться не будет.
Еще раз коротко: ЭОС - это про направление движения электричества по сердечной мышце.
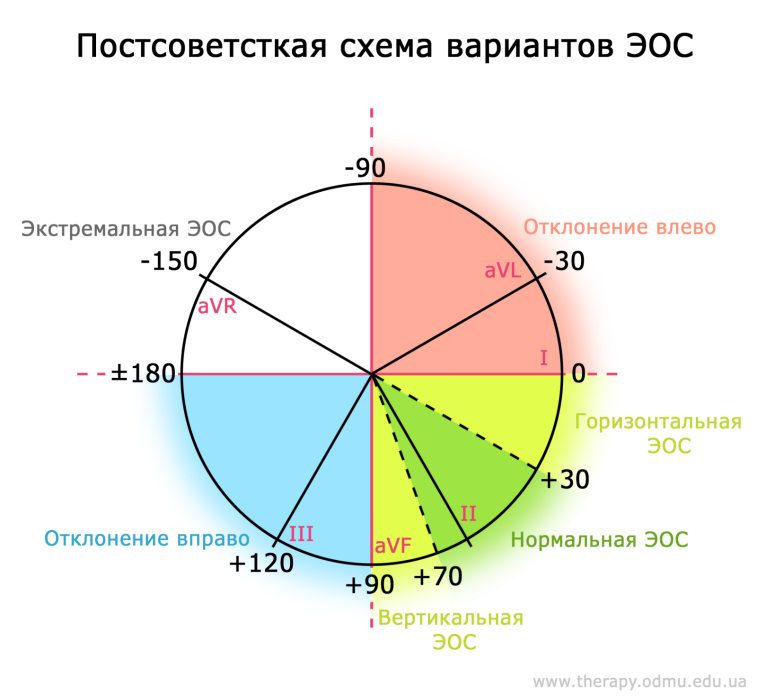
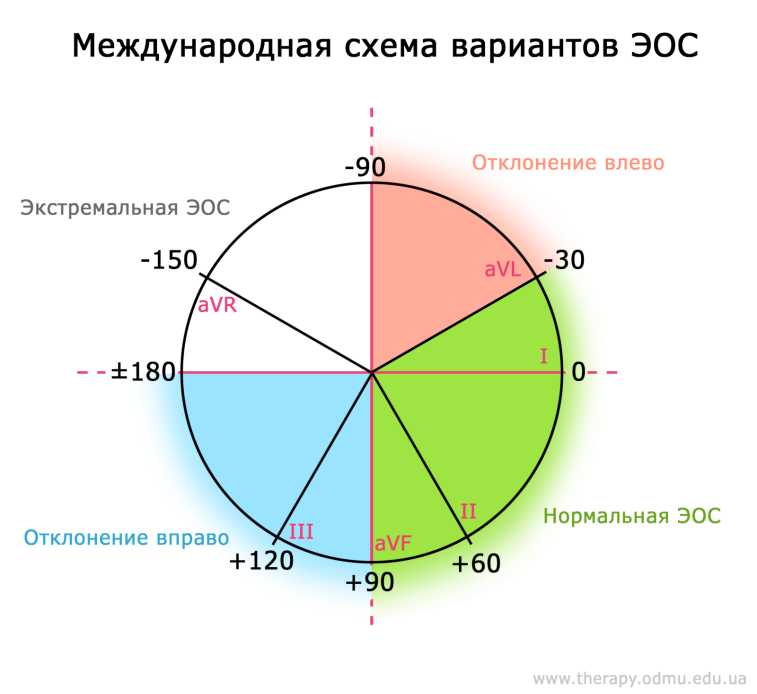
Какой бывает ЭОС?
В постсоветстких странах система отведений несколько отличается от международно принятой: существуют т.н. "горизонтальная" и "вертикальная" ЭОС, которые в других странах отдельно не выделяются и входят в понятие нормы.
Наглядно разница видна на этих двух схемах:
| 
|
Как видно, сейчас выделяют четыре положения ЭОС:
§ Нормальная (от -30о до 90о)
§ Отклонение влево (от -30о до -90о)
§ Отклонение вправо (от 90о до 180о)
§ Экстремальная правая ЭОС (от -90о до 180о)
Пример №1
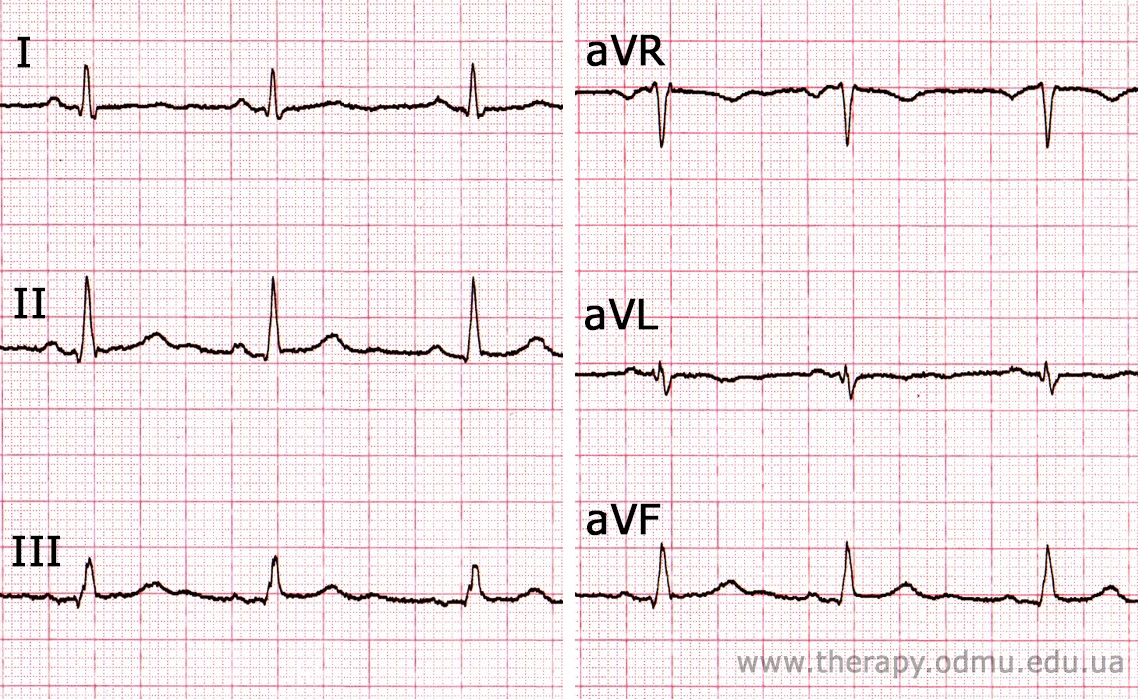
§ Мы видим, что самый высокий зубец R в отведении II . Это значит, что волна в основном шла в его сторону.
§ Самый глубокий S в отведении aVR - значит, волна шла ОТ него.
§ В отведении aVL комплекс QRS состоит из одинакового положительного R и отрицательного S - это значит, что волна сначала приближалась кэтому электроду, а потом от него удалялась (прошла мимо).
§ Электрическая ось данного пациента совпадает со II отведением. Глядя на диаграмму выше делаем вывод, что ось - нормальная, угол α = 60°
Пример №2
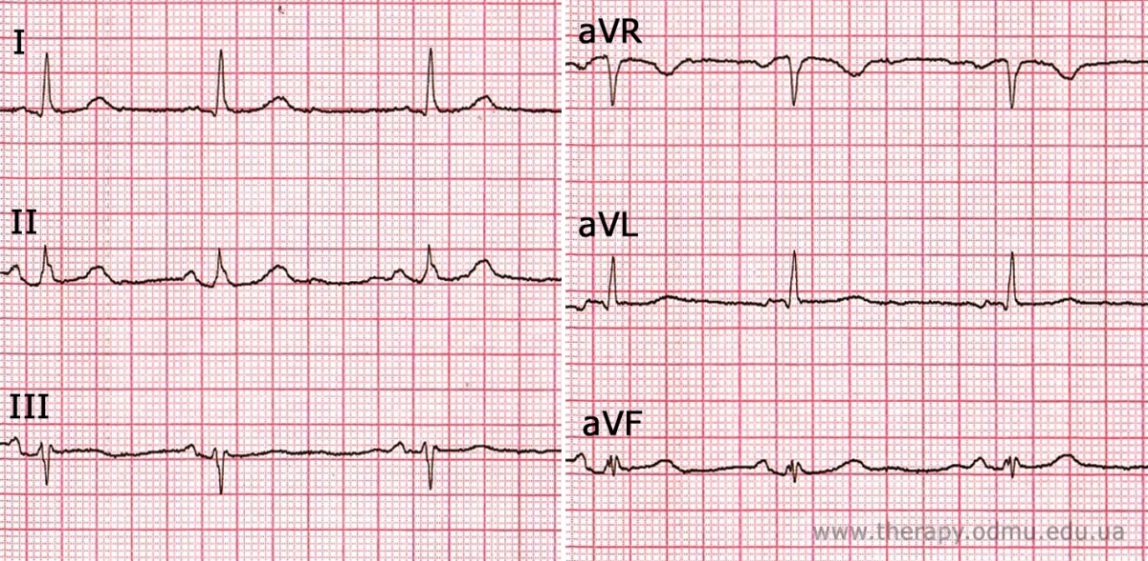
§ Самый высокий зубец R в отведении I (к нему шла волна деполяризации)
§ Самый глубокий S в отведениях III и aVR - значит, волна шла от них.
§ Изоэлектрический комплекс QRS виден в отведении aVF - значит волна деполяризации прошла поперек этого отведения.
§ Подведем итог: электрическая волна прошла от правых отведений (III, aVR) к I отведению, пройдя поперек отведения aVF. Смотрим на диаграмму чуть выше и определяем ось, как горизонтальную (по-новому: нормальную), угол α = 0°
Описание установки.
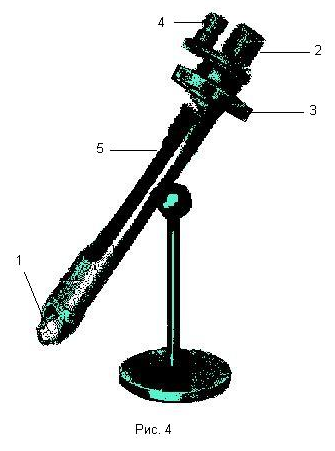
В работе используется поляриметр портативный П-161, внешний вид которого изображен на рисунке 4. Оптическая схема прибора изображена на рисунке 5.
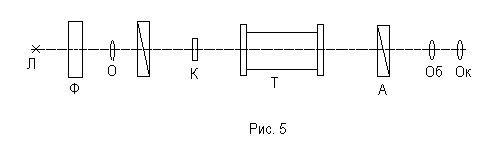
Источником света в поляриметре может являться лампа накаливания или солнечный свет, направляемый при помощи зеркальца. Свет от источника падает на светофильтр Ф и объектив О. Полученный монохроматический свет проходит через поляризатор П, кювету Т с раствором и анализатор А. В качестве анализатора и поляризатора в приборе используются поляроиды. После анализатора свет проходит через объектив Об и окуляр Ок зрительной трубы сахариметра, которая служит для визуального наблюдения поля зрения.
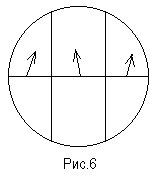 Вследствие адаптации глаза визуально трудно оценивать абсолютную освещенность. В то же время легко сравнивать освещенность различных частей поля зрения. Для разделения поля зрения на части в сахариметре непосредственно за поляризатором расположена тонкая кварцевая пластинка К, через которую проходит средняя часть пучка поляризованного света, вышедшего из поляризатора.
Вследствие адаптации глаза визуально трудно оценивать абсолютную освещенность. В то же время легко сравнивать освещенность различных частей поля зрения. Для разделения поля зрения на части в сахариметре непосредственно за поляризатором расположена тонкая кварцевая пластинка К, через которую проходит средняя часть пучка поляризованного света, вышедшего из поляризатора.
В результате введения кварцевой пластинки поле зрения поляриметра оказывается разделенным на три части. Средняя часть освещается светом, прошедшим через поляризатор, кварцевую пластинку и анализатор, а две крайние части поля зрения - светом, прошедшим через поляризатор и анализатор. Так как кварц является оптически активным веществом, то после прохождения поляризованного света через пластинку его плоскость поляризации поворачивается на некоторый угол (рис.6).
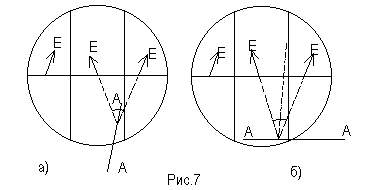 Поворачивая анализатор, можно получить равномерно освещенное поле зрения. Это будет происходить при двух положениях анализатора: 1) плоскость АА анализатора совпадает с биссектрисой угла между направлениями колебаний в средней и крайней частях поля зрения (рис.7,а); 2) плоскость анализатора перпендикулярна биссектрисе угла между направлениями колебаний (рис.7,б). В одном случае яркость поля зрения будет больше, в другой - меньше. При работе с сахариметром следует уравнивать части поля зрения при меньшей яркости.
Поворачивая анализатор, можно получить равномерно освещенное поле зрения. Это будет происходить при двух положениях анализатора: 1) плоскость АА анализатора совпадает с биссектрисой угла между направлениями колебаний в средней и крайней частях поля зрения (рис.7,а); 2) плоскость анализатора перпендикулярна биссектрисе угла между направлениями колебаний (рис.7,б). В одном случае яркость поля зрения будет больше, в другой - меньше. При работе с сахариметром следует уравнивать части поля зрения при меньшей яркости.
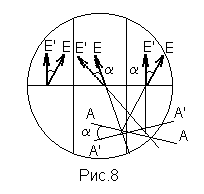 Если установить анализатор на равную освещенность всех частей поля зрения, а затем поместить между поляризатором и анализатором трубку с раствором сахара, то равенство яркостей средней и крайней частей поля зрения нарушиться. Это происходит вследствие того, что во всех частях поля зрения плоскость колебаний светового вектора повернется на один и тот же угол a (рис.8). Для восстановления равенства освещенностей необходимо повернуть анализатор на этот же угол a, равный углу поворота плоскости поляризации света при прохождении им раствора сахара.
Если установить анализатор на равную освещенность всех частей поля зрения, а затем поместить между поляризатором и анализатором трубку с раствором сахара, то равенство яркостей средней и крайней частей поля зрения нарушиться. Это происходит вследствие того, что во всех частях поля зрения плоскость колебаний светового вектора повернется на один и тот же угол a (рис.8). Для восстановления равенства освещенностей необходимо повернуть анализатор на этот же угол a, равный углу поворота плоскости поляризации света при прохождении им раствора сахара.
Преломление света — явление, при котором луч света, переходя из одной среды в другую, изменяет направление на границе этих сред.
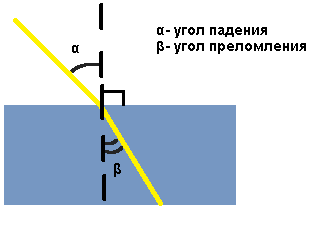
Преломление света происходит по следующему закону: Падающий и преломленный лучи и перпендикуляр, проведенный к границе раздела двух сред в точке падения луча, лежат в одной плоскости. Отношение синуса угла падения к синусу угла преломления есть величина постоянная для двух сред: 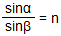 , гдеα — угол падения, β — угол преломления, n — постоянная величина, не зависящая от угла падения.
, гдеα — угол падения, β — угол преломления, n — постоянная величина, не зависящая от угла падения.
При изменении угла падения изменяется и угол преломления. Чем больше угол падения, тем больше угол преломления. Если свет идет из среды оптически менее плотной в более плотную среду, то угол преломления всегда меньше угла падения: β < α.Луч света, направленный перпендикулярно к границе раздела двух сред, проходит из одной среды в другую без преломления.
абсолютный показатель преломления вещества — величина, равная отношению фазовых скоростей света (электромагнитных волн) в вакууме и в данной среде n=c/v Величина n, входящая в закон преломления, называется относительным показателем преломления для пары сред.
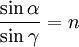
|
Величина n есть относительный показатель преломления среды В по отношению к среде А, а n' = 1/n есть относительный показатель преломления среды А по отношению к среде В. Эта величина при прочих равных условиях больше единицы при переходе луча из среды более плотной в среду менее плотную, и меньше единицы при переходе луча из среды менее плотной в среду более плотную (например, из газа или из вакуума в жидкость или твердое тело). Есть исключения из этого правила, и потому принято называть среду оптически более или менее плотной, чем другая. Луч, падающий из безвоздушного пространства на поверхность какой-нибудь среды В, преломляется сильнее, чем при падении на нее из другой среды А; показатель преломления луча, падающего на среду из безвоздушного пространства, называется его абсолютным показателем преломления .
(Абсолютный - относительно вакуума. Относительный - относительно любого другого вещества (того же воздуха, например). Относительный показатель двух веществ есть отношение их абсолютных показателей. )
Полное внутреннее отражение — внутреннее отражение, при условии, что угол падения превосходит некоторый критический угол. При этом падающая волна отражается полностью, и значение коэффициента отражения превосходит его самые большие значения для полированных поверхностей. Коэффициент отражения при полном внутреннем отражении не зависит от длины волны.
В оптике это явление наблюдается для широкого спектра электромагнитного излучения, включая рентгеновский диапазон.
В геометрической оптике явление объясняется в рамках закона Снелла. Учитывая, что угол преломления не может превышать 90°, получаем, что при угле падения, синус которого больше отношения меньшего показателя преломления к большему показателю, электромагнитная волна должна полностью отражаться в первую среду.
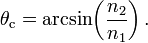
В соответствии с волновой теорией явления, электромагнитная волна всё же проникает во вторую среду — там распространяется так называемая «неоднородная волна», которая экспоненциально затухает и энергию с собой не уносит. Характерная глубина проникновения неоднородной волны во вторую среду порядка длины волны.
Вследствие взаимодействия электромагнитной волны со средой, изменяется скорость её распространения. Эта зависимость имеет вид: v=c/n, где n= 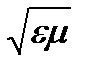 -абсолютный показатель преломления вещества, v – скорость света в среде, а с – скорость света в вакууме. При переходе света через границу раздела двух сред, скорость распространения света в которых различна, происходит изменение его направления. Это явление называется преломлением или рефракцией света. Явление рефракции света легло в основу метода определения концентрации разбавленных растворов по эмпирической зависимости между показателем преломления и концентрацией раствора.
-абсолютный показатель преломления вещества, v – скорость света в среде, а с – скорость света в вакууме. При переходе света через границу раздела двух сред, скорость распространения света в которых различна, происходит изменение его направления. Это явление называется преломлением или рефракцией света. Явление рефракции света легло в основу метода определения концентрации разбавленных растворов по эмпирической зависимости между показателем преломления и концентрацией раствора.
Относительный показатель преломления сред
n21=n2/n1
где n2 и n1 - абсолютные показатели преломления сред.
При переходе света из среды с меньшим показателем преломления (оптически менее плотная среда) в среду с большим показателем преломления (оптически более плотная среда) угол падения луча больше угла преломления. Если луч падает на границу раздела сред под наибольшим возможным углом i=p/2 (луч скользит вдоль границы раздела сред), то он будет преломляться под углом r<p/2. Этот угол является наибольшим углом преломления для данных сред и называется предельным углом преломления. Из закона преломления света следует
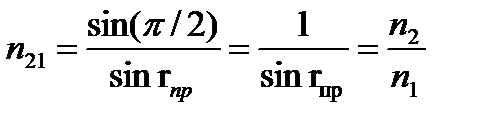 , откуда
, откуда
sin rпр=n1/n2
Если свет переходит из оптически более плотной среды в оптически менее плотную, то угол преломления больше угла падения. При некотором угле падения i луча угол преломления равен p/2, т.е. преломлённый луч скользит вдоль границы раздела сред. При дальнейшем увеличении угла падения преломление не происходит, весь падающий свет отражается от границы раздела сред (полное отражение). Угол i называется предельным углом полного отражения и обозначается iпр. Так как
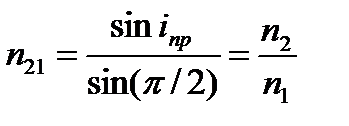 ,
,
то
sin iпр=n 2/n 1
Таким образом, предельный угол преломления и предельный угол полного отражения для данных сред зависят от их показателей преломления. Это нашло применение в приборах для измерения показателя преломления веществ - рефрактометрах, используемых при определении чистоты воды, концентрации общего белка сыворотки крови, для идентификации различных веществ и так далее.
Описание установки
Основной частью рефрактометра являются две прямоугольные призмы 1 и 2, сделанные из одного и того же сорта стекла (рис.1,а). Призмы соприкасаются гипотенузными гранями, между которыми имеется зазор около 0,1 мм. Между призмами помещают каплю жидкости, показатель преломления которой требуется определить. Луч света от источника 3 направляется на боковую грань верхней призмы и, преломившись, попадает на гипотенузную грань АВ. Поверхность АВ матовая, поэтому свет рассеивается и, пройдя через исследуемую жидкость, падает на грань CD нижней призмы под различными углами от 0 до 90°. Если показатель преломления жидкости меньше показателя преломления стекла, то лучи света входят в призму 2 в пределах от 0 до rпр. Пространство внутри этого угла будет освещенным, а вне его – тёмным. Таким образом, поле зрения, видимое в зрительную трубу, разделено на две части: тёмную и светлую. Положение границы раздела света и тени определяется предельным углом преломления, зависящим от показателя преломления исследуемой жидкости.
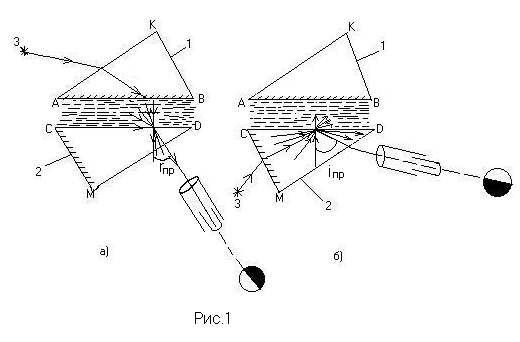
Если исследуемая жидкость имеет большой показатель поглощения (мутная, окрашенная жидкость), то во избежание потерь энергии при прохождении света через жидкость измерения проводят в отраженном свете. Ход лучей в рефрактометре в этом случае показан на рис.1,б. Луч света от источника проходит через матовую боковую грань СМ нижней призмы 2. При этом свет рассеивается и падает на гипотенузную грань CD, соприкасающуюся с исследуемой жидкостью, под всевозможными углами от 0 до 90°. Если жидкость оптически менее плотная, чем стекло, из которого изготовлена призма, то лучи, падающие под углами, большими iпр, будут испытывать полное отражение и выходить через вторую боковую грань нижней призмы в зрительную трубу. Поле зрения, видимое в зрительную трубу, так же как и в первом случае, окажется разделенным на светлую и тёмную части. Положение границы раздела в данном случае определяется предельным углом полного отражения, также зависящем от показателя преломления исследуемой жидкости.
С помощью этого прибора можно исследовать вещества, показатель преломления которых меньше показателя преломления стекла измерительных призм. Оптическая система рефрактометра изображена на рис. 2.
В рефрактометре используется источник 3 белого света. Вследствие дисперсии при прохождении светом призм 1 и 2 граница света и тени оказывается окрашенной. Во избежание этого перед объективом зрительной трубы помещают компенсатор 4. Он состоит из двух одинаковых призм, каждая из которых склеена из трех призм, обладающих различным показателем преломления. Призмы подбирают так, чтобы монохроматический луч с длиной волны l = 589,3 мкм. (длина волны желтой линии натрия) не испытывал после прохождения компенсатора отклонения. Лучи с другими длинами волн отклоняются призмами в различных направлениях. Перемещая призмы компенсатора с помощью специальной рукоятки, добиваются того, чтобы граница света и темноты стала возможно более чёткой.
Лучи света, пройдя компенсатор, попадают в объектив 6 зрительной трубы. Изображение границы раздела свет – тень рассматривается в окуляр 7 зрительной трубы. Одновременно в окуляр рассматривается шкала 8. Так как предельный угол преломления и предельный угол полного отражения зависят от показателя преломления жидкости, то на шкале рефрактометра сразу нанесены значения этого показателя преломления.
Оптическая система рефрактометра содержит также поворотную призму 5. Она позволяет расположить ось зрительной трубы перпендикулярно призмам 1 и 2, что делает наблюдение более удобным.
В общей фокальной плоскости объектива и окуляра зрительной трубы помещают стеклянную пластинку, на которую нанесена визирная линия (или крест, образованный тонкими нитями). Перемещением зрительной трубы добиваются совпадения визирной линии с границей свет – тень и по шкале определяют показатель преломления исследуемой жидкости. В некоторых современных рефрактометрах зрительная труба укрепляется неподвижно, а система измерительных призм может поворачиваться.
Описание установки.
Для выполнения работы могут быть использованы универсальный фотоэлектроколориметр типа КФК-2 или более современный КФК-2МП.
В основу устройства фотоэлектроколориметра положен принцип уравнивания двух световых потоков путем изменения одного из них с помощью диафрагмы с переменным отверстием.
Если диафрагмы одинаково освещены и в одинаковой мере раскрыты, то яркость обеих половин поля зрения будут одинаковы. Если же при равенстве яркостей на пути одного светового потока, например первого, поместить объект, частично поглощающий свет, то фотометрическое равенство нарушится, так как правая половина поля зрения станет менее яркой. Для того чтобы уровнять яркость полей, необходимо ослабить интенсивность пучка, уменьшив отверстие диафрагмы, через которую он проходит.
При измерении концентрации вещества в растворах на пути одного из пучков света ставится стеклянная кювета с исследуемым раствором. Для того чтобы учесть поглощение света растворителем, на пути второго пучка ставится такая же кювета с чистым растворителем. Количество жидкостей в обеих кюветах должно быть одинаковым.
Для проведения измерений в монохроматическом свете прибор снабжён одиннадцатью светофильтрами, расположенными в револьверном диске. При повороте диска номера светофильтров появляются в его окошечке. Восемь светофильтров делят видимую область спектра на примерно равные участки шириной в среднем 40 нм. Остальные обладают более широкой полосой пропускания и делят видимую область на три части: красную, зелёную и синюю. Светофильтры характеризуются эффективной длиной волны lэф, соответствующей максимуму коэффициента пропускания для данного светофильтра. Эффективные длины волн светофильтров и их маркировка приведены в следующей таблице.
Гамма-излучение
Взаимодействие гамма-квантов с веществом может сопровождаться фотоэффектом,комптоновскимрассеянием и образованиемэлектрон-позитронных пар. Вид эффекта зависит от энергии гамма-кванта:
Ек = hν – Еи, (1)
где: h –постоянная Планка;ν –частота излучения;Еи – энергия ионизации соответствующей атомной оболочки (энергия связи выбитого электрона из атома).
Фотоэффектвозникает приЕ = 10 эВ–1 МэВ, то есть при относительно малых значениях энергий. В этом случае вся энергия гамма-кванта передается орбитальному электрону, и он выбивается из орбиты (рис.3).
Выбитый электрон называется фотоэлектроном. В результате его отрыва в атоме появляется свободный уровень, который заполняется одним из наружных электронов. При этом, либо испускается вторичное мягкое характеристическое излучение (процесс флюоресценции), либо энергия передается одному из электронов, который покидает атом (электрон Оже). Флюоресцентное излучение наблюдают в материалах с большим атомным номером. В материалах с низким атомным номером преобладает образование электронов Оже. Вероятность фотоэффекта увеличивается с ростом атомного номера материала и уменьшается с ростом энергии фотона.
С ростом энергии гамма-квантов явление фотоэффекта становится все меньше, а при энергии 100–200 кэВ начинает преобладать Комптон эффект.
Комптоновским рассеиваниемназывается процесс взаимодействия фотонного излучения с веществом, в котором фотон в результате упругого столкновения с орбитальным электроном теряет часть своей энергии и изменяет направление своего первоначального движения, а из атома выбивается электрон отдачи (комптоновский электрон) (рис.4).
Энергия комптоновского электрона равна:
Е = hν – hν\(2)
Образование электронно-позитронных пар. Если энергия гамма кванта превышает1,02 МэВ, то он поглощается ядром, а из последнего одновременно вылетают электрон и позитрон (рис.5). Таким образом, гамма кванты способны косвенно ионизировать вещество. Возникшей паре передается вся энергия гамма кванта за вычетом энергии покоя пары, равной 1,022 МэВ.
Следует отметить, что позитрон нестабилен в присутствии электронов среды. Он быстро исчезает за счет аннигиляции с одним из электронов. В этом случае испускается 2 фотона с энергией по 0,511 МэВ.
Рассмотрим, проникающую способность гамма-квантов.
Как уже отмечалось, гамма-квант образуется при переходе ядра в более низкие энергетические состояния. Не имея массы, они не могут замедляться в среде, а лишь поглощаются или рассеиваются.

 При прохождении через вещество их энергия не меняется, но уменьшается интенсивность излучения по следующему закону (рис.6):
При прохождении через вещество их энергия не меняется, но уменьшается интенсивность излучения по следующему закону (рис.6):
I = Iо е–- µх (3)
где: I = Еγn/t; n/t– число гамма-квантов, падающих на единицу поверхности в единицу времени (плотность потока гамма-квантов);m–коэффициент поглощения;х– толщина поглотителя (вещества), см;Iо –интенсивность квантов до прохождения поглотителя, МэВ/с.
В формуле (3) величину µ можно найти в таблицах, но она не несет прямой информации о степени поглощения гамма лучей веществом.
В практических расчетах удобно пользоваться и такой табличной величиной, как «толщина слоя половинного ослабления». Толщина слоя половинного ослабления – это такая толщина слоя материала, проходя которую интенсивность излучения гамма-квантов уменьшается в 2 раза. Запишем уравнение (3) в виде:
Iо /I = е– µх(4)
Полагая Iо/I = 2 и логарифмируя правую и левую части уравнения (4), получим:ln2 =md, d = 0,693/m.
Тогда, формула (4) примет вид:
I = Iо е– 0,693х/d(5)
Толщина слоя половинного ослабления d берется из таблиц, но если они отсутствуют, то эта величина может быть вычислена приближенно по плотности материалаρ:
d = 13/r,(6)
где: 13 см– слой воды, ослабляющий гамма-излучение в 2 раза;r –плотность материала, г/см3. Для некоторых материалов величиныdпредставлены в таблицах.
γ≈
γ■≈
I0
I

 х
х
 Рис.6. К оценке ослабления гамма-излучений веществом
Рис.6. К оценке ослабления гамма-излучений веществом
Выражение (5) можно преобразовать следующим образом:
Косл = I0/I = ехр (0,693х/d),(7)
где Косл – коэффициент ослабления гамма-излучения проходящего через преграду толщинойх и значением слоя половинного ослабления для данного материалаd(рис.6). Выражение (7) можно упростить, полагая, что 0,693 =Ln2, получим:
Косл = 2х/d (8)
Расчеты показывают, что проникающая способность гамма-излучения в воздухе – десятки и сотни метров, в твердых телах – многие сантиметры, в биологической ткани человека часть гамма-квантов проходят через человека насквозь, другие поглощаются.
Биофизика как наука. Современные достижения биофизики и их значение для биологии и медицины.
Биофизика — это наука, изучающая физические свойства биологических объектов, а также физические и физико-химические закономерности, лежащие в основе их функционирования. Непосредственным объектом исследования биофизики является живая материя на разных уровнях ее организации (начиная от молекулярного и кончая организменным или даже популяционным). Биофизика, наряду с биохимией, биоорганической химией, молекулярной биологией, иммунологией и рядом других дисциплин, входит в состав комплекса наук, объединяемых под названием физико-химической биологии.
Изучение организма человека и процессов, связанных с нарушением его жизнедеятельности, является предметом медицинской биофизики. Достижения медицинской биофизики углубляют наши знания о механизмах возникновения и развития болезней, способствуют разработке новых лабораторных методов диагностики, совершенствованию методов лечения.
Значительна роль медицинской биофизики в изучении физико-химических основ патологии; методы биофизики, приспособленные к условиям клинических лабораторий, все шире используются в целях диагностики, а также для оценки эффективности проводимого лечения. Биофизические методы применяют при создании новых лекарственных средств как на стадии их первичного отбора, так и на стадии выяснения механизма действия. Наконец, на основе методов и теоретических достижений биофизики создаются новые и совершенствуются традиционные способы лечения больных.
В современной биофизике условно выделяют квантовую биофизику, молекулярную биофизику, биофизику клетки и биофизику сложных систем (т. е. биофизику органов и тканей, системы организм—среда и т. д.).
Квантовая биофизика.
Раздел науки, изучающий электронное строение биологических молекул, электронные переходы в них, процессы превращения энергии возбужденных состояний молекул в другие виды энергии. Предметом исследования квантовой биофизики являются: строение электронных энергетических уровней молекул; донорно-акцепторные свойства биологических молекул; поглощение света веществом и люминесценция с точки зрения электронных переходов; механизм свободнорадикальных процессов и свойства свободных радикалов; химические реакции электронно-возбужденных молекул; химические свойства фотопродуктов; механизм хемилюминесценции.
Молекулярная биофизика.
Молекулярная биофизика — раздел биофизики, изучающий физическую структуру биологически важных молекул, прежде всего белков и нуклеиновых кислот, и физические процессы, лежащие в основе их функционирования с целью выяснения связи физической структуры и свойств с выполняемой ими в организме функцией. Является междисциплинарной наукой, включающей в себя методы и исследования в таких областях, как биохимия, генетика, физика, компьютерное моделирование и так далее. Молекулярная биофизика изучает структурные химические формулы, длины всех связей и углы между ними, распределение электрических зарядов на поверхности молекул, подвижность отдельных участков молекул и изменчивость структуры молекул в зависимости от параметров среды.
Биофизика мембран.
Достижения в области биофизики мембран имеют первостепенное значение для медицинской науки и практики, т. к. известно, что нарушение работы мембранных систем является одной из причин нарушений функционирования клеток. Именно нарушение функционирования мембранных структур приводит к биологической смерти клеток при гипоксии, интоксикациях, механических повреждениях тканей, отморожениях и ожогах, действии ультрафиолетового и, возможно, ионизирующего излучения. Доказано изменение свойств мембранных структур при многих заболеваниях нервной системы, при развитии атеросклероза и ишемической болезни сердца, при гепатитах, заболеваниях почек.
Нарушения функционирования мембран могут быть следствием изменения активности работы мембранных ферментов, деятельности мембранных рецепторов или ионных каналов.
Знание механизмов функционирования и нарушения работы мембранных систем дает возможность применения большого числа разнообразных лекарственных средств (адренолитиков, местноанестезирующих препаратов, наркотических средств, антигистаминных препаратов и др.) для коррекции работы мембранных каналов и рецепторов.
Биофизика клеточной подвижности и мышечного сокращения.
Изучение механизмов мышечного сокращения с позиций биофизики и данные электронной микроскопии позволили разработать молекулярные модели мышечного сокращения. Открытие опорного и сократительного аппарата в немышечных клетках расширило наши представления о распространенности явлений подвижности клеточных элементов. Изучение механизма движения протоплазмы у простейших показало, что движение в клетке может быть вызвано не только механохимическими явлениями, но и хемоосмотическими процессами. Функционирование внутриклеточных сократительных белков в
тромбоцитах составляет важное звено в сложном процессе агрегации этих клеток, приводящем к образованию тромбов. Представляется перспективным поиск лекарственных средств, действующих на регуляцию процессов агрегации тромбоцитов. Результаты биофизического исследования мышечного сокращения легли в основу разработки системы количественных показателей сократимости миокарда с целью диагностики заболеваний сердца.
Биофизика сложных систем.
Одним из крупнейших достижений биофизики сложных систем является открытие и расшифровка механизма возникновения автоколебаний в биологических системах, а также распространение автоволн в возбудимых средах, к к-рым, в частности, относится миокард. Химической моделью таких процессов может служить реакция Белоусова — Жаботинского (окисление лимонной кислоты в присутствии бромата и ионов церия). Появлением спонтанных центров зарождения автоволн были объяснены особенности движения протоплазмы у ряда организмов, движение крови в сосудах мышечного типа и возникновение сердечных аритмий. Понимание механизма автоколебательных и автоволновых процессов — необходимый этап в решении ряда проблем хронобиологии, в эффективном использовании принципов хрономедицины, в разработке методов лечения аритмий сердца и других заболеваний, связанных со спонтанной возбудимостью нервных и мышечных тканей. Многие свойства автоколебаний и автоволн в сложных системах изучают в основном с помощью методов математического моделирования.
Широкое использование ЭВМ позволило существенно приблизиться к решению ряда проблем, связанных с математическим моделированием биологических процессов в норме и при патологии. Одной из них является проблема возникновения электрических полей в организме. В частности, формальное описание электрического поля сердца не давало достаточного основания для понимания связи изменений в работе отдельных мышечных клеток и ткани в целом с изменениями в отводимых от тела потенциалах при электрокардиографии. Создание математической модели, позволяющей анализировать ЭКГ человека с учетом морфологических, цитологических и физиологических параметров миокарда, дает возможность сопоставлять изменения электрокардиограммы с определенными нарушениями функционирования миокарда, а следовательно, улучшать существующие методы диагностики и лечения сердечно-сосудистых заболеваний. Значительные успехи достигнуты в исследованиях физических основ природы электроэнцефалограммы. Анализ электрической активности отдельных нейронов и факт наличия синхронизации активности многих нейронов позволили дать физико-математическое описание статистических особенностей ЭЭГ, что расширяет возможности ее диагностического применения.
Одной из традиционных отраслей биофизики, используемых в медицине, является биомеханика. Результаты изучения механических свойств мышц, связок, костей, стенок кровеносных сосудов, легких оказались важными для травматологии, кардиологии, пульмонологии и других областей медицины. Все большее внимание уделяется молекулярным и клеточным основам механических свойств биологических объектов, включая синовиальную жидкость, кожу, мышцы, альвеолы легких и стенки сосудов. Достигнут прогресс в исследовании реологических свойств форменных элементов крови, биологических мембран, в изучении закономерностей движения крови по мелким кровеносным сосудам, в т. ч. по капиллярам, и т. д. Методы биомеханики легли в основу новых методов клинической диагностики, напр, определение упругости стенок сосудов и ее изменения при атеросклерозе. Измерение механических характеристик синовиальной жидкости, крови, кожи, кровеносных сосудов, костей и других тканей и органов используется для оценки их патологических изменений. Изучение способности эритроцитов к межклеточному взаимодействию, заключающемуся в образовании агрегатов и наблюдающемуся при ишемической болезни сердца, нарушениях кровообращения в различных органах, позволило разработать клинические способы коррекции механических свойств крови.
Изучение гемодинамики получило дальнейшее развитие благодаря широкому внедрению методов математического и физического моделирования гемодинамиче-ских процессов. Результаты изучения гемодинамики создали теоретическую основу для конструирования аппаратов искусственного кровообращения, протезирования сердца, его клапанов, кровеносных сосудов, создания искусственной почки, для разработки способов замены крови с помощью искусственных переносчиков кислорода и т. д.
Исследования гемодинамических процессов, механизма генерации и распространения пульсовой волны, кар-диогенных смещений тела привели к разработке новых бескровных физических способов изучения функционирования сердца, нашедших применение в клинической практике.
Появились новые методы диагностики, к-рые иногда объединяют под названием «медицинская инженерия» или медицинская физика. В частности, происходит активная разработка и широкое внедрение в медицину диагностических методов, основанных на использовании различных видов излучений. К их числу относятся рентгеновская компьютерная томография и томография с использованием ядерного магнитного резонанса, ультразвуковая визуализация внутренних органов и тканей, эхолокация, широко используемые в офтальмологии, кардиологии, акушерстве; ультразвуковая допплеровская спектроскопия, применяемая для измерения скорости кровотока, и многие другие методы.
Дата: 2019-04-22, просмотров: 633.