Винер не разделял теорию трех генов Фишера – Рейса, оставаясь последо-вательным приверженцем концепции одного гена. Не признал он и кроссин-говер. Действительно, при наличии одного гена кроссинговер маловероятен. Убежденность, с которой Винер отстаивал свои взгляды, побуждает критически отнестись к рассмотрению этого вопроса.
Легко приняв на веру подкупающую простотой теорию Фишера – Рейса, мало кто из специалистов, кроме Винера, подверг ее серьезной перепроверке. Прокоп и Гёллер в фундаментальном труде «Группы крови человека» [90] пи-шут, что Винер критиковал теорию кроссинговера, неоднократно проверяя ее по таблицам популяционно-генетических исследований и не находя в них под-тверждения ожидаемого кроссинговера. Напротив, некоторые позиции противо-речили теории Фишера.
По мнению Рейса [544], на кроссинговер указывали лишь единичные наблю-дения, из которых трудно было сделать однозначное заключение о существова-нии этого феномена.
концепции Типпетт также нет места кроссинговеру. Трудно ожидать пере-креста двух расположенных рядом тесно сцепленных локусов. При таких усло-виях более вероятны делеции, мутации и конверсии.
Как и любое теоретическое построение, рассмотренные выше 3 генетиче-ские теории – это лишь предположения, попытки систематизировать, объяснить экспериментальные данные, исходя из представлений того времени.
Сегодня можно высказать суждение (ни в коей мере не подвергая сомнению теорию Фишера), что порядок расположения генов RH может соответствовать последовательности D – E – C и это ничего не меняет на фенотипическом уров-не. Кроссинговер (если он в системе Rh происходит) может дать такие же соче-тания антигенов при последовательности генов D – E – C, как и при последова-тельности D – C – E (см. рис. 4.2). Последовательность генов C – D – E также ничего не меняет в Rh-фенотипе человека, если допустить возможность выбо-рочной конверсии генетического материала при мейозе.
Гаплотип cDe встречается в 10–13 раз чаще у негроидов, чем у европеоидов (42,3 и 3,2 % соответственно [108]). Если бы гаплотип cDe являлся результатом кроссинговера, как полагал Фишер, то частота гаплотипов Cde, cdE, CDE и CdE также должна быть существенно выше у негроидов, чем у европеоидов. Однако
и действительности частота указанных гаплотипов у представителей этих двух рас приблизительно одинакова. Тем не менее идея Фишера о том, что редкие га-плотипы образуются посредством кроссинговера частых гаплотипов, признает-ся всеми исследователями как весьма элегантная, и если кроссинговер не был до сих пор убедительно доказан, то он и не был полностью опровергнут.
Теоретические построения Типпетт, при всей их оригинальности, также не могут рассматриваться как истина в последней инстанции. В них много допу-щений. Не ясно: почему чаще образуются антитела анти-С, анти-с, анти-Е и
166
анти-е, чем антитела анти-се, анти-Се, анти-сЕ и анти-СЕ, хотя обе группы ан-тител стимулированы, как полагает Типпетт, одним полипептидом? Почему так часто образуются несепарируемые анти-DC-антитела, если антигены D и C находятся на разных полипептидах? Почему чаще вырабатываются анти-DЕ-антитела, чем анти-Е, но реже, чем анти-DC? Винер объяснял это существова-
нием двух агглютиногенов: Rhо' (DC) и Rhо" (DE), которые встречаются с раз-ной частотой. По мнению Фишера, это объясняется тем, что гены D и Е, а зна-чит и антигены D и Е, дальше отстоят друг от друга, чем D и С, поэтому веро-ятность образования анти-DC-антител выше, чем анти-DE. С позиций концеп-ции Типпетт образование комбинированных антител анти-DC и анти-DE мож-но объяснить, допустив, что эпитопы Rh мозаично переплетены на поверхности эритроцитов в виде близкорасположенных пар DC и DE.
Концепция двух генов пока еще осмысливается иммуносерологами, привык-шими оперировать категориями Винера, Фишера и Рейса. Если антитела анти-се, анти-Се, анти-сЕ и анти-СЕ определяют продукты гена RHСЕ, то почему ан-титела анти-DC и анти-DE не могут свидетельствовать о существовании гена RHDCE с аллелями DC и DE, подобно тому, как считал Винер?
Пока ответом на этот вопрос служит открытие двух разных протеинов, несу-щих антигены D и СЕ. Однако не исключено, что в ближайшем будущем могут быть найдены протеины, несущие одновременно специфичность D и С, D и Е. Гибридные гены DC–D / D–DC, продуцирующие необычные иммунодоминант-ные протеины, известны. Вместе с тем следует признать, что теория двух генов представляет несомненный прогресс в иммуносерологии и весьма перспектив-на для дальнейших молекулярно-биологических изысканий.
Как справедливо указывают Issitt, Anstee [374], дискуссия относительно трех генетических теорий системы резус далека от завершения. Однако эта дискуссия не содержит антагонистических противоречий. Как первая, так и вторая, и тре-тья теории не противоречат практике и вполне устраивают иммуносерологов, су-дебных медиков, генетиков и других специалистов. Различаясь по форме, эти кон-цепции никак не сказываются на интерпретации результатов фенотипирования при использовании конкретных тестовых реагентов. В этих теориях практически все позиции общие, за исключением количества детерминирующих генов.
Номенклатура Фишера – Рейса не противоречит номенклатуре Винера, так как опирается на одни и те же факты (обе исследовательские группы, Рейса в Англии и Винера в Америке, обменивались найденными сыворотками и сопо-ставляли полученные результаты). Концепция Типпетт никаких изменений в су-ществующую номенклатуру не внесла.
30. все-таки, может быть, более всех прав Винер, и наблюдаемое разнообра-зие фенотипов резус, несмотря на национальные и расовые особенности, обе-спечивается одним геном? Многочисленных воздействий на дистанции «фор-мирование гена → ген → готовый продукт» в виде кроссинговера, конверсии, мутации, делеции, пространственного взаимовлияния генов друг на друга и
167
всего, что может воздействовать на кодирующую ДНК и синтез полипептидов, более чем достаточно, чтобы обеспечить существующее разнообразие. Вряд ли для этого нужно 3, а тем более 50 генов, достаточно одного. Теория Типпетт по-строена в унисон теории Винера. Она по сути представляет собой возврат от те-ории трех генов или более к теории одного гена.
Наследование
Rh-антигены передаются индивиду по наследству в виде двух гаплотипов: одного – от отца, другого – от матери. Как и при наследовании других групповых признаков, у детей не может быть Rh-антигенов, отсутствующих у родителей.
Часто лекторы для закрепления знаний у слушателей спрашивают: «Могут ли родиться резус-положительные дети у резус-отрицательных родителей?»
36 получив, как правило, отрицательный ответ, продолжают вопрос: «А у резус-положительных родителей – резус-отрицательные дети?». На второй вопрос так-же нередко сдедует отрицательный ответ: «Не могут!». Это неверно. У резус-положительных родителей могут родиться резус-отрицательные дети, если оба родителя гетерозиготны – D / d × D / d.
Ниже приведена удобная для использования схема, позволяющая отразить возможные Rh-фенотипы детей, которые можно ожидать у конкретной супру-жеской пары (рис. 4.3).
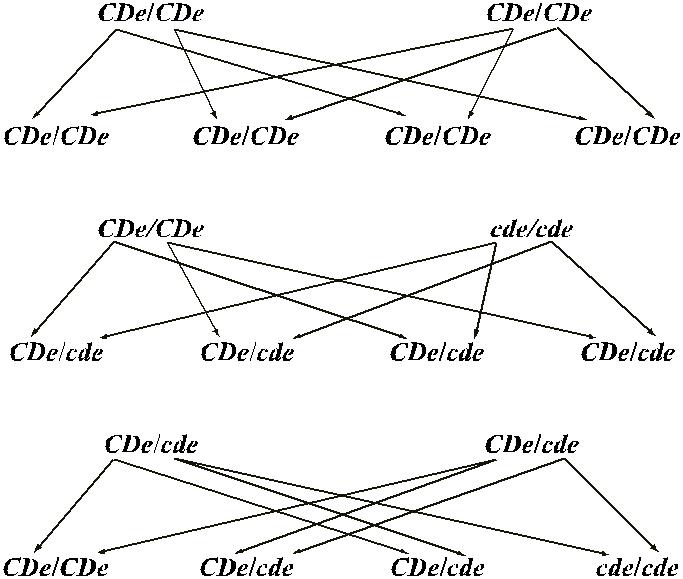
Рис. 4.3. Варианты наследования антигенов Rh.
9. супругов CDe / CDe × CDe / CDe все дети будут гомозиготы CDe / CDe. В се-мье, где один из родителей CDe / CDe, а другой – cde / cde, все дети будут гетерози-готы CDe / cde. Когда родители гетерозиготы CDe / cde × CDe / cde, 75 % детей будут резус-положительными, 25 % – резус-отрицательными; из всех детей 25 % будут гомозиготы CDe / CDe, 50 % – гетерозиготы CDe / cde и 25 % – гомозиготы cde / cde.
168
Используя такую схему, легко просчитать другие варианты фенотипа и генотипа детей или, наоборот, по фенотипу детей установить предполагаемый фенотип ро-дителей.
Если фенотип можно определить с помощью сывороток, то генотип, как пра-вило, устанавливают априори, исходя из частоты встречаемости того или иного фенотипа. В большинстве случаев данные совпадают. Например, генотип резус-отрицательного человека в 97,5 % случаев соответствует cde / cde или cde/–; ге-нотип человека Cde почти в 100 % случаев Сde / cde, генотип человека CDe при-мерно в 30 % случаев CDe / cde, а в 16 % случаев – CDe / CDe. Высока (более 90 %) вероятность того, что человек с фенотипом cDe имеет генотип cDe / cde, а не cDe / cDe, поскольку гаплотип cde встречается значительно чаще, чем гапло-тип cDe. Более точно установить RH-генотип человека можно на основании се-мейного исследования, т. е. установления Rh-фенотипа родителей, братьев, се-стер и детей (близких родственников, желательно 3–4 поколений).
[36] высокой точностью генотип может быть установлен с помощью набо-ра сывороток анти-се, анти-Се, анти-сЕ и анти-СЕ. Например, человек с фено-типом СсDEe может иметь генотип cDE / Cde или CDE / cde. При обоих геноти-пах эритроциты будут реагировать одинаково с сыворотками анти-D, анти-С, анти-Е, анти-с и анти-Е, а с сыворотками анти-се, анти-Се, анти-сЕ и анти-СЕ – по-разному. При генотипе cDE / Cde агглютинация эритроцитов произой-дет с сыворотками анти-Се и анти-сЕ, но не произойдет с сыворотками анти-се
и анти-СЕ (см. табл. 4.7). При генотипе CDE / cde, наоборот, эритроциты будут реагировать с сыворотками анти-се и анти-СЕ, но не будут реагировать с сыво-ротками анти-Се и анти-сЕ. Точно также эритроциты CcEe с генотипом Cde / cdE будут реагировать с сыворотками анти-Се и анти-сЕ, а у человека с генотипом CdE / cde – только с сыворотками анти-се и анти-СЕ.
Наиболее точно (но не в 100 % случаев) генотип устанавливают, анали-зируя сам ген. Для этого исследуют ДНК человека с помощью молекулярно-
биологических методов (см. ДНК-типирование Rh-антигенов).
Эффекты транс и цис
Гаплотип одной хромосомы может влиять на экспрессию антигенов, коди-руемых гаплотипом другой хромосомы (транс). Наиболее выраженный эффект позиции транс прослеживается у лиц с генотипами Cde / cDe и Cde / CDe, при ко-торых ген С, находящийся в положении транс по отношению к гену D, приво-дит к продукции слабого D-антигена (см. раздел Фенотип D u), в то время как ген D кодирует нормальную экспрессию D-антигена, когда в положении транс находится ген с [202, 204]. На рис. 4.4 представлены результаты исследования 2 семей, у членов которых этот эффект четко прослеживался.
В дяди и племянницы, имевших генотип Cde / cDe, экспрессия антигена D была снижена вдвое по сравнению с тремя другими членами семей, генотип ко-торых был CDe / сde (215, 219 и 444, 440, 450 соответственно).
169
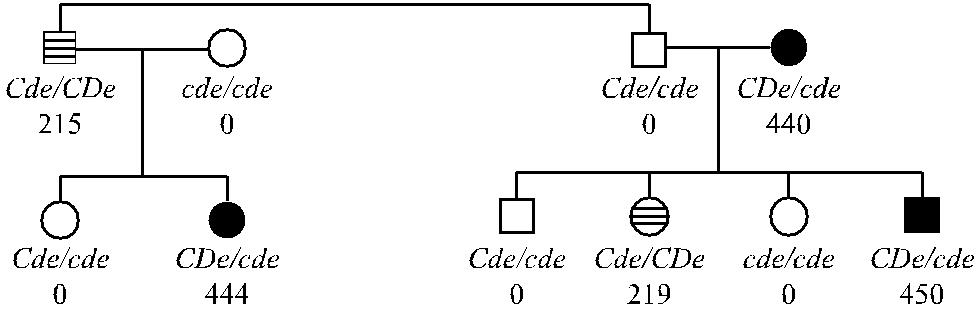
Рис. 4.4. Влияние гаплотипа Cde на экспрессию антигена D (по Ceppellini и соавт. [202]). Черные фигуры – нормальное количество D-антигена, заштрихованные – сниженное количество D-антигена, белые – отсутствие D-антигена. Цифры означают количество D-антигена в эритроцитах, выраженное в виде среднего титра 6 сывороток анти-D, титрованных с данными эритроцитами.
Race и Sanger [544], исследуя специально отобранные сыворотки анти-С и анти-Е, установили, что гены С и Е в позиции цис и транс влияют на экспрессию одноименных антигенов (табл. 4.9). Ген Е в позиции цис угнетает продукцию ан-тигена С, а ген С в позиции транс – продукцию антигена Е. Впоследствии авто-ры несколько изменили свои взгляды, придя к выводу, что эффект супрессии мо-жет быть обусловлен не взаимодействием генов, а наличием в указанных, заранее отобранных сыворотках примеси других антител. Некоторые сыворотки анти-С содержат фракцию анти-Се-антител, поэтому сильнее реагируют с эритроцита-ми лиц Cde / cdE и Cde / СDe, имеющими оба антигена (С и Се). Сыворотки анти-Е иногда содержат анти-СЕ-антитела, за счет чего сильнее реагируют с эритроцита-ми лиц CdE / cde и CDE / cDe, несущими антигены Е и СЕ.
Однако для того чтобы полностью исключить взаимовлияние генов С и Е в разных генетических комбинациях, нет достаточных оснований. На других мо-делях эффект транс и цис в той или иной мере проявляется.
Таблица 4.9
Дата: 2019-02-24, просмотров: 249.